Diversity and Environmental Specificity of Fungal Isolates from Soils and Cereal Grains in Western Kenya
-
Edward Mwakichwa Mwatabu

Department of Seed, Crop and Horticultural Science, School of Agriculture and Biotechnology, University of Eldoret, P.O. Box, 1125-30100, Eldoret, Kenya
Javan Omondi WereDepartment of Seed, Crop and Horticultural Science, School of Agriculture and Biotechnology, University of Eldoret, P.O. Box, 1125-30100, Eldoret, Kenya
Josiah Chemulanga ChiveuDepartment of Seed, Crop and Horticultural Science, School of Agriculture and Biotechnology, University of Eldoret, P.O. Box, 1125-30100, Eldoret, Kenya
Evans Ochieng OumaDepartment of Agriculture and Environmental Studies, School of science, Agriculture and Environmental Studies, Rongo University, P.O. Box, 103-40404, Rongo, Kenya
Meshack ObonyoDepartment of Biochemistry and Molecular Biology, Egerton University, Njoro, Kenya
| Received 22 Sep, 2023 |
Accepted 16 Mar, 2024 |
Published 31 Mar, 2024 |
Background and Objective: Mycotoxin contamination in maize and groundnuts has undermined Kenya's health care system for decades. This study aimed to determine the diversity of mycotoxin fungi in maize, groundnut and soils of Western Kenya and farmers' awareness of on-field mycotoxin mitigation measures. Materials and Methods: Infected maize, groundnut and soil samples were collected from Homa Bay, Migori, Siaya and Busia Counties. Semi-structured questionnaires were used to collect socio-economic data on mycotoxin awareness and mitigation measures used by farmers. Pure fungal isolates were obtained for diversity assessment on PDA and incubated at 25-27°C. A light microscope at ×400 magnification was used for the morphological identification of spores and mycelia, while species were identified using plant pathology reference books and journals. Results: Thirty-five diverse fungal isolates were obtained from all samples. The genus Aspergillus was the highest in terms of isolation frequency, with 14 diverse pathotypes, followed by Penicillium (8 isolates), Fusarium (4 isolates) and the rest (9 isolates). Busia County had the most diverse number of isolates, while Siaya had the least. Among sample categories, most fungi were obtained from soil samples (30 isolates) while maize and groundnuts recorded 19 and 9 isolates respectively. While 22 isolates were specific to counties, 13 were environmentally non-specific. Farmers in Western Kenya had partial knowledge of best pre- and post-harvest mycotoxin mitigation practices. Conclusion: Mycotoxin-producing fungi were the most dominant fungi in maize, groundnut and soils of Busia, Siaya, Homa Bay and Migori counties of Western Kenya. These fungal species were highly specific to the environments.
| Copyright © 2024 Mwatabu et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Mycotoxins contaminate various food substances and agricultural products worldwide, posing serious health risks to humans and animals1,2. The presence of mycotoxin in significant quantities may induce acute to chronic health effects such as endocrine disruption in humans and animals3. Human exposure to mycotoxins may result from consumption of plant-derived foods with toxin adulteration. Additionally, exposure may be instigated by metabolites in animal products such as meat and eggs or exposure to air and dust-containing toxins4. Over 65% of the Kenyan population is exposed to chronic levels of mycotoxin poisoning5-6. Common mycotoxin-producing fungi, such as Aspergillus flavus species, thrive well in substrates of grains such as maize and groundnuts which are the most widely consumed crops7-9. Maize-groundnut intercrops are very popular in the Western region of Kenya10. This is because groundnut crop is known to fix large amounts of nitrogen and is considered an ideal option for rotation and intercropping with maize fields to boost yields11-13. The cereal-legume intercrop system is highly practiced by farmers as a mean of diversification to mitigate climate change and for managing striga weed menace which is a major constraint to crop productivity in this region.
However, a bulk of farmers, producers and consumers have little knowledge of best farming practices against mycotoxin accumulation in grain/field or the experience in dealing with mycotoxin contamination in foodstuff and grain14-16. Therefore, people in these regions are at a high risk of aflatoxin contamination. Inappropriate pre-and post-harvest procedures augment colonisation by mycotoxin-producing fungi of field crops and post-harvest produce17-20. Inadequate drying and improper storage coupled with high temperature and humidity levels are essential contributing factors to the accumulation of mycotoxins up to unacceptable levels in developing countries21-23. The risk is further aggravated as these farmers chiefly sell their surplus through informal markets which are highly unregulated5,24. Such farmers' practices are risk factors for mycotoxin contamination. Enhancing farmer, producer and consumer knowledge on mycotoxin mitigation measures right from the early stages of production to storage is highly desired as a means to mitigate the problem of mycotoxin contamination. However, sufficient farmers' knowledge on causes and sustainable management of these mycotoxin fungi on soils, maize and groundnuts in Western Kenya is still lacking several studies conducted in Western Kenya such as those of Were et al.25 and Lewis et al.26 in Homa bay and Bungoma counties detected the presence of aflatoxin in these regions. Others such as Lewis et al.26 and Nelson et al.27 determined aflatoxin levels in sample grains. Additionally, information on the occurrence, diversity and farmer knowledge of mycotoxin fungi in Busia County among others is still inadequate. This implies that the diversity of mycotoxin-producing fungi and non-toxigenic species in the region is a critical research gap and a vital step in identifying potential bio-control agents. Therefore, this study was established to assess the diversity of fungi in maize and groundnuts in Western Kenya and the levels of awareness of the causes and mitigations of aflatoxins in maize and groundnut grains among smallholders in Western Kenya.
MATERIALS AND METHODS
Study location and environmental characteristics: The study duration was from September 2019 to April 2022. Surveys were conducted in three sub-counties (sub-regions) of four counties of Western Kenya namely Busia, Siaya, Homa Bay and Migori. The four study regions were selected on the basis that maize and groundnuts are among the major food crops grown by the majority of farmers there. The four sampling sites have different agroecological conditions, ambient temperatures and annual rainfall. The climatic data of these four regions is unique and favours mycotoxin fungi establishment and growth (Table 1).
Sample collection, sample size determination and sampling technique: The study areas were grouped into clusters based on regional boundaries32. Sub-counties were selected randomly from each cluster to represent a sampling hub. Three significant towns serving sub-county headquarters within the four counties were selected as sampling hubs. The sampling sites include villages representing different AEZs within a 20 km radius of the sampling hub. The GPS-detecting applications and administrative boundaries were used for site location and mapping. Two hundred and fifty grams each of maize, groundnut grains (from the previous season) and soil samples were collected from selected farmers with a long history of maize-groundnut intercropping systems. The process generated information on the incidence and prevalence of aflatoxin under different farming practices. During the sample collection, a total of 46 maize and groundnut farmers, 11 (Busia), 5 (Siaya), 15 (Homabay) and 15 (Migori), were selected and interviewed on grain storage methods and level of awareness on mycotoxin fungi with reference to commonly recommended mitigation measures using a semi-structured questionnaire33. Data on farmer level of awareness and commonly practiced mitigation measures in the region were subjected to analysis using descriptive statistics and presented in bar graphs with standard error bars using Microsoft Excel 365.
| Table 1: | Weather details of sampling sites of Western Kenya | |||
| Country | Annual average temperature (°C) |
Annual rainfall (mm) |
Highest humidity (%) |
Source |
| Busia | 21.8 |
2291 |
82.59-May |
Atsiaya et al.28 |
| Siaya | 21.4 |
2154 |
82.74-April |
Maina et al.29 |
| Homa Bay | 21.7 |
1331 |
71.5-May |
Ogenga et al.30 |
| Migori | 21 |
1522 |
76.88-April |
Olela et al.31 |
Isolation and culture purification of grain and soil fungi: Two hundred and fifty grams of grain (maize and groundnut) and soil samples which were aseptically collected from each farmer as described in section 2.2 above were used for fungal extraction. Isolation and purification of the mycotoxin fungi from diseased groundnut, maize seed and soil was done according to Salano et al.34 and Owino et al.35. Five grains per sample were surface sterilised in 1% sodium hypochlorite (NaOCl) for 2 min, rinsed thrice in sterile distilled water, then plated on sterile PDA media and incubated at 27°C in the dark. After 5 days, a single conidium representing each collection was transferred to a freshly prepared PDA and set at similar conditions. Sub-cultures were made from the emerging colonies and pure cultures were obtained for subsequent studies36. Soil samples were passed through a 2 mm mesh sieve to remove crop residues and 1 gram of the sample was suspended in 9 mL sterile distilled water. The suspension was serially diluted to 10–3 and a 200 μL aliquot of the dilution was uniformly spread in duplicates on PDA37. The plates were incubated for five days, after which colonies of Aspergillus and other fungi were counted and the number of Colony Forming Units per gram (CFU/g) of soil was calculated according to Okayo et al.38:
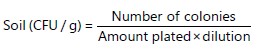 |
Fungal diversity assessment and identification: Mycotoxin fungal isolates from all regions of the study were subjected to morphological descriptive assays and identification was done using various pathology reference journals such as39-43. Additionally, the assessment of diversity was based on observations using a biological light microscope (model MOMxianweijing2933 from Beijing, China) at ×400 magnification as well as in the petri-dishes to classify and identify isolates with respect to spore shape and mycelial growth characteristics such as mycelial colour, (front and reverse side colour) and texture. The Isolates were also grouped based on agro-ecological zones (counties) of isolation to document those specific to certain regions and those found across all the regions considered in the study.
RESULTS
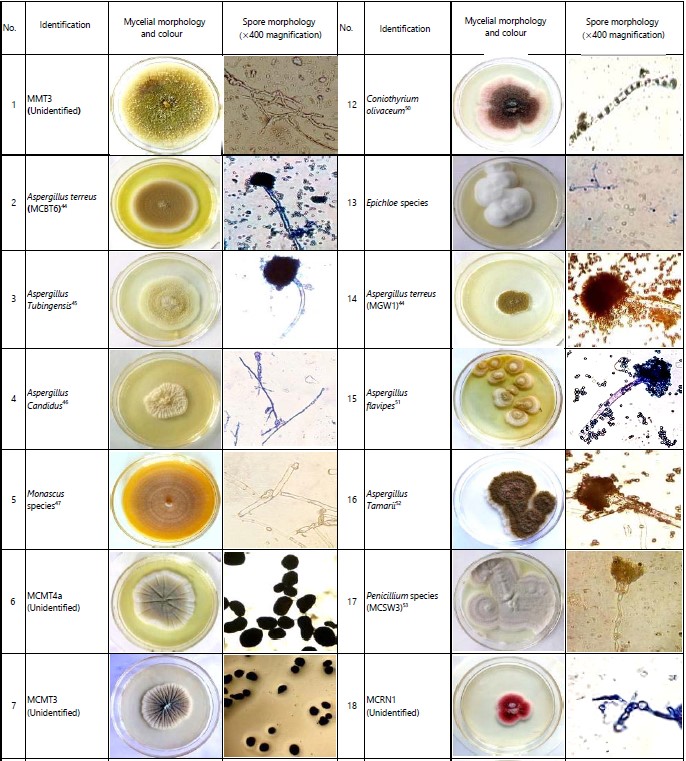
Morphological diversity and identification of soil and grain fungi in Western Kenya: Diversity assessment and identification assay recorded a total of 35 diverse fungal isolates from groundnut grain, maize grain and soil samples across the four counties of Western Kenya. Out of the 35 diverse isolates, the genus Aspergillus was the highest in terms of frequency with 14 diverse pathotypes, followed by the genus Penicillium (8 isolates) and Fusarium (4 isolates), respectively (Table 2 and 3). Also, even within a fungal species, significant morphological differences were observed physically in plates and at the microscopy level in terms of mycelial colour, substrate colour and spore shape. While Busia County had the highest number of diverse isolates in all sample types, Siaya County had the lowest number of isolates.
| Table 2: | Occurrence and isolation frequency of environmental/region-specific fungal isolates | |||
Counties in Western Kenya |
|||||||||||||
Busia |
Siaya |
Homa Bay |
Migori |
||||||||||
| Identified and unidentified fungus (region and sample specific) |
M |
G |
S |
M |
G |
S |
M |
G |
S |
M |
G |
S |
Frequency of isolation |
| MMT3 (Unidentified) | + |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
| Aspergillus terreus (MCBT6) | - |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
| Aspergillus tubingensis | - |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
| Aspergillus candidus | - |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
| Monascus species | - |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
| MCMT4a (Unidentified) | - |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
| MCMT3 (Unidentified) | - |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
| MCMBT3 (Unidentified) | - |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
| Arthrinium sacchari | - |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
1 |
| Coniothyrium olivaceum | - |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
1 |
| Epichloe species | - |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
1 |
| Aspergillus terreus (MGW1) | - |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
1 |
| Aspergillus flavipes | - |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
1 |
| Aspergillus tamarii | - |
- |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
1 |
| MCHB2 (Unidentified) | - |
- |
- |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
1 |
| MCRN1 (Unidentified) | - |
- |
- |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
1 |
| Aspergillus species (MSE2) | - |
- |
- |
- |
- |
- |
- |
- |
- |
+ |
+ |
+ |
3 |
| Aspergillus parasiticus | - |
- |
- |
- |
- |
- |
- |
- |
- |
+ |
- |
- |
1 |
| Aspergillus oryzae | - |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
+ |
1 |
| Penicillium species (MCSW3) | - |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
+ |
1 |
| Phialemoniopsis endophytica | - |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
+ |
1 |
| Aspergillus species (MCSW1) | - |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
+ |
1 |
| Number of fungi per sample | 1 |
0 |
7 |
0 |
0 |
3 |
2 |
1 |
2 |
2 |
1 |
5 |
24 |
| Number of fungi per region (Country) |
8 |
3 |
5 |
8 |
|||||||||
| M: Maize grain, G: Groundnut grain and S: Soil sample | |||||||||||||
| Table 3: | Occurrence and isolation frequency of environmental non-specific fungal isolates | |||
Counties in Western Kenya |
|||||||||||||
Busia |
Siaya |
Homa Bay |
Migori |
||||||||||
| Isolate identity | M |
G |
S |
M |
G |
S |
M |
G |
S |
M |
G |
S |
Frequency of isolation |
| Aspergillus flavus (MUG5) | + |
- |
- |
+ |
- |
+ |
+ |
- |
- |
+ |
- |
- |
5 |
| Aspergillus niger | + |
+ |
+ |
+ |
- |
+ |
- |
- |
+ |
+ |
- |
+ |
8 |
| Penicillium aurantiogriseum | + |
+ |
- |
- |
+ |
- |
- |
+ |
+ |
- |
+ |
- |
6 |
| Penicillium species (MMT2) | + |
+ |
+ |
- |
- |
- |
- |
- |
- |
- |
+ |
- |
4 |
| Fusarium oxysporum (MGW5) | + |
- |
+ |
+ |
+ |
- |
+ |
- |
- |
- |
- |
- |
5 |
| Fusarium proliferatum | + |
- |
- |
- |
- |
+ |
- |
+ |
- |
- |
- |
- |
3 |
| Aspergillus nomius | - |
- |
+ |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
2 |
| Aspergillus flavus (GMT3) | - |
+ |
- |
- |
+ |
- |
- |
+ |
- |
- |
- |
- |
3 |
| Fusarium species (MCBT1) | - |
+ |
+ |
- |
- |
+ |
- |
- |
+ |
- |
- |
+ |
5 |
| Penicillium chrysogenum | - |
- |
+ |
- |
- |
- |
- |
+ |
- |
- |
- |
+ |
3 |
| MCMT4b (Unidentified) | - |
- |
- |
- |
- |
+ |
- |
- |
+ |
- |
- |
- |
2 |
| Trichoderma harzianum | - |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
+ |
2 |
| Biatriospora species | - |
- |
- |
- |
- |
- |
- |
- |
+ |
- |
- |
+ |
2 |
| Number of fungi per sample | 6 |
5 |
6 |
3 |
3 |
5 |
3 |
4 |
6 |
2 |
2 |
5 |
50 |
| Number of fungi per region (Country) |
17 |
11 |
13 |
9 |
|||||||||
| M: Maize grain, G: Groundnut grain and S: Soil sample | |||||||||||||
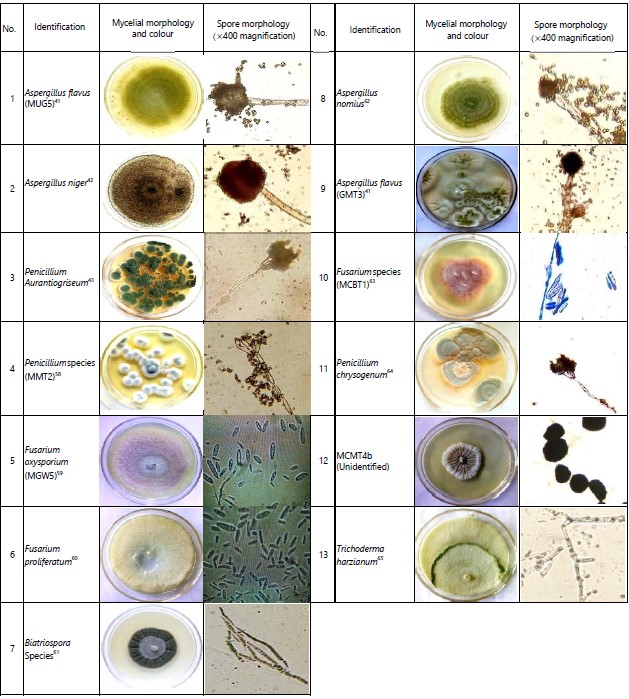
Aspergillus flavus (MUG5) was the most common isolate in maize samples, as it was detected in all sample regions. This isolate, however, was not detected in groundnut samples. Fusarium oxysporum (MGW5) and Aspergillus niger (MBT2) were detected in three out of four counties. Aspergillus flavus (GMT3) was the most common isolate among groundnut samples and was detected in Busia, Siaya and Homa Bay counties. Alternately, Aspergillus niger (MBT2) was the most common isolate in soil samples, as was detected in all counties of the sample regions (Table 2). Thirty-one isolates were characterised to the generic level, while four isolates remained unidentified.
|
Environmental specificity and non-specificity of fungal isolates in Western Kenya: Generally, the majority of the fungal species isolated from maize, groundnut and soil samples were specific to the environments in which the samples were collected (Fig. 1). Twenty-two diverse fungal isolates were specific to one region/county (Fig. 1). Among the isolates, 10 were identified as Aspergillus species, with only one isolate from the Penicillium genus. Most of these isolates were isolated from Busia County samples (Table 2). However, out of the 9, Busia County-specific isolates, 8 were isolated from soil samples, one isolate obtained from maize samples and none detected in groundnuts. Siaya County recorded the least number of unique isolates (3) detected in soil samples only. Additionally, out of the 22 region-specific isolates, only four were specific to maize samples, while only one was specific to groundnut samples (Table 2).
On the other hand, thirteen diverse fungal isolates were non-specific to their environments (Fig. 2). Among these isolates, 12 were detected in Homa Bay County and 10 in Busia County, while Siaya and Migori counties recorded 8 diverse fungal isolates each (Table 3). However, in terms of isolation frequency, these isolates were most frequently detected in Busia County followed by Homa Bay County and lastly in Migori County.
|

|
|
Colony Forming Units (CFUs) of soil-inhabiting fungal species in Western Kenya: At the county level, there were no significant differences in CFU/g between Busia, Homa Bay, Migori and Siaya Counties. In terms of variation from one CFU to the other, Busia, Homa Bay and Migori were the highest. However, Siaya had the least variation in terms of CFU, though with the highest mean above 17 CFU/g. Additionally, with respect to sample distribution (whiskers), Busia County recorded the highest number of colony-forming units per unit gram (>30 CFU/g), while Homa Bay had the lowest (<5 CFU/g). Also, Siaya had the highest mean, while Homa Bay County had the lowest mean of CFU/g in soil samples (Fig. 3).
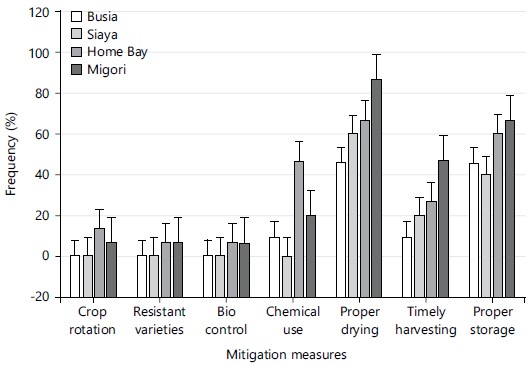
Farmer awareness levels on mitigation of mycotoxins: Farmer assessments on mycotoxin management revealed low awareness levels on mitigation measures. Proper drying and proper storage of grains were the most known mitigation measures against mycotoxin accumulation as they recorded at least 40% awareness levels across the four counties. However, other mitigation measures such as the use of resistant varieties, use of chemicals and crop rotation scored less than 20% across the four counties (Fig. 4).
DISCUSSION
Fungal diversity and environmental specificity: The fungal diversity assessment study revealed that the soils of Western Kenya are enriched with a comprehensive genetic bank of soil and seed-inhabiting fungi. Therefore, mycotoxin accumulation remains a major threat to safe maize and groundnut products in western Kenya. This finding was mainly because the most predominant fungal isolates from the collected samples belonged to the three major mycotoxin-producing genera: Aspergillus, Penicillium and Fusarium66. The great diversity displayed in mycotoxin and infectious fungal species isolated portrays their abundance and possible widespread distribution through evolution36,67. For example, Aspergillus flavus was the most frequently detected fungus and occurred in two different pathotypes implying that the fungus has a great evolutionary potential. In addition to confirming its ubiquitous nature, the findings may also imply that mycotoxin accumulation will remain a challenge in the region for decades if not adequately alleviated68-70. High diversity in mycotoxin-producing fungi is an imminent management threat. However, such diversity also sets the stage towards the identification and development of biological mycotoxin control measures where non-toxigenic strains are identified and used.
In addition to diversity within a genus, inter-genera diversity was the highest. The presence of many fungal isolates from other genera insinuates the possibility of identifying other bio-control agents against mycotoxin accumulation in grains and soils. Aspergillus flavus (MUG5) was detected in both maize and soil samples. This finding implies that the fungus is both a soil and seed-borne pathogen. These findings compare well with those of Monda et al.71 who isolated Aspergillus flavus from soil samples obtained from maize fields and suggested that the most effective way of managing this pathogen is by reducing its accumulation in the soil environment as well as on grain. Further analysis of fungal diversity disclosed that the majority of fungal isolates were obtained from the soil samples (Table 2 and 3). By comparing the counties, the magnitude of fungal diversity in soil samples positively correlated with diversity levels in maize and grain samples (Table 3). For example, Busia County also had the most diverse isolates of all the sample types. However, this scenario was more evident in mycotoxin fungi than in the rest of the isolates. This could be associated with the nature of their spores. For example, Aspergillus and Penicillium species produce many air-borne spores and hence are transmitted directly from the soil to seed in the field72,73.
Besides soils, maize grains were more infected by fungal isolates obtained in the study than groundnuts. This occurrence could be due to a pathogen or race specificity. The division of Forma specialis of these mycotoxin fungi into different races explains why fungi with similar morphologies may have dissimilar infection rates on host plants74. However, this study cannot fully ascertain this phenomenon because the fungal isolates were not positively identified to the race level. Also, the grains assessed were only samples from the study region; hence the study is bound by the limitation of the sampling process.
For sample-specificity, 22 out of the total 35 fungal isolates were environmentally specific. This finding was expected due to the eminent environmental differences known to exist among the four counties. Further, the occurrence and prevalence of toxigenic and non-toxigenic fungi is highly dependent on the environmental conditions75. Busia County had the most diverse number of isolates, while Siaya County had the least diverse number of isolates (Table 2 and 3) probably due to variations in awareness levels on mitigation of fungal accumulation and variations in environmental conditions76. Respondents from Busia County were least aware of post-harvest mitigation measures against the accumulation of aflatoxin fungi. Such unawareness is a gateway to practices such as improper drying of grains providing sufficient moisture for fungal growth and proliferation and this may contribute to the diversity and abundance of fungi77.
Farmer awareness levels and mycotoxin fungal diversity: Farmers in the sampled regions had little knowledge of mitigation measures against mycotoxin accumulation in grains (Fig. 4). These findings add tremendously to the scientific body of knowledge by revealing the current status of aflatoxin awareness levels in Western Kenya regions a subject that has been scarcely documented78-81. This knowledge has a ripple effect on fungal diversity because farmers are unlikely to deliberately use pre-and post-harvest practices that inhibit mycotoxin accumulation. For example, a common storage practice such as the use of gunny bags favors moisture build-up promoting fungal growth when placed on floors21,82,83. Low awareness levels on mycotoxin mitigation encourage practices that expose the grains to conditions that favors penetration and multiplication of mycotoxin-producing fungi84-86. Farmers are also unlikely to consistently practice programs such as crop rotation which reduce inoculum load and insect pest infestations87. Understanding these aspects by farmers is likely to reduce the inoculum load of most toxigenic fungi. Risk exposure indicators include knowledge of the causes of aflatoxin contamination and possible mitigations5,6. This knowledge is also mostly shared in farmer discussion forums. However, such initiatives are not practiced consistently in Western Kenya.
Implications, applications, recommendations and limitations of the study: The study findings are vital since they prompt further ground-truthing on the status of mycotoxin risk and vacuity of sustainable solutions in Western Kenya. The occurrence nature of identified fungal species in terms of diversity and environmental specificity is valuable information that can guide relevant geographical scope for identified bio-controls. Critical limitations of this study include the small geographical scope and number of plant species assessed. Additionally, molecular species identification was not conducted. Therefore, genetic chractersization of fungal species identified and in-vitro studies to identify species with the capacity to suppress mycotoxin fungi in Western Kenya is recommended.
CONCLUSION AND RECOMMENDATIONS
The study findings demonstrated that Aspergillus species are the most dominant fungi in maize, groundnut and soils of Busia, Siaya, Homa Bay and Migori Counties of Western Kenya. Although most fungal isolates in the region are sample and regional specific, mycotoxin fungi are non-specific to environments and samples. This study also revealed that farmers in Western Kenya have little awareness of appropriate mycotoxin mitigation measures. Therefore, there is a need for characterization and identification of mycotoxin-producing fungi in the region for sustainable management. Additionally, deliberate efforts need to be made to enhance information dissemination to farmers on on-farm mycotoxin management strategies.
SIGNIFICANCE STATEMENT
This study aimed to assess the diversity of mycotoxin fungi in maize, groundnut and soils in Western Kenya and farmers' awareness of mycotoxin mitigation measures. Thirty-five diverse pure cultures of fungal isolates were obtained from sampled cereal grains and soils of Western Kenya. The genus Aspergillus had the highest isolation frequency, with fourteen diverse pathotypes, followed by Penicillium (eight isolates), Fusarium (four isolates) and the rest (nine isolates). Busia County had the most diverse number of isolates, while Siaya had the least. While twenty two isolates were specific to counties, thirteen were environmentally non-specific. Farmers in Western Kenya had partial knowledge of best pre- and post-harvest mycotoxin mitigation practices.
REFERENCES
- de Ruyck, K., M. de Boevre, I. Huybrechts and S. de Saeger, 2015. Dietary mycotoxins, co-exposure, and carcinogenesis in humans: Short review. Mutat. Res. Rev. Mutat. Res., 766: 32-41.
- Danso, J.K., E.A. Osekre, G.P. Opit, N. Manu and P. Armstrong et al., 2018. Post-harvest insect infestation and mycotoxin levels in maize markets in the Middle Belt of Ghana. J. Stored Prod. Res., 77: 9-15.
- Awuchi, C.G., I.C. Owuamanam, C.C. Ogueke and T. Hannington, 2020. The impacts of mycotoxins on the proximate composition and functional properties of grains. Eur. Acad. Res., 8: 1024-1071.
- Zain, M.E., 2011. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc., 15: 129-144.
- Mutegi, C.K., P.J. Cotty and R. Bandyopadhyay, 2018. Prevalence and mitigation of aflatoxins in Kenya (1960-to date). World Mycotoxin J., 11: 341-357.
- Mutiga, S.K., J.M. Mutuku, V. Koskei, J.K. Gitau and F. Ng’ang’a et al., 2021. Multiple mycotoxins in Kenyan rice. Toxins, 13.
- Agbetiameh, D., A. Ortega-Beltran, R.T. Awuah, J. Atehnkeng and M.S. Islam et al., 2019. Potential of atoxigenic Aspergillus flavus vegetative compatibility groups associated with maize and groundnut in Ghana as biocontrol agents for aflatoxin management. Front. Microbiol., 10.
- Omara, T., A.K. Kiprop, P. Wangila, A.P. Wacoo and S. Kagoya et al., 2021. The scourge of aflatoxins in Kenya: a 60-year review (1960 to 2020). J. Food Qual., 2020.
- Senghor, L.A., A. Ortega-Beltran, J. Atehnkeng, K.A. Callicott, P.J. Cotty and R. Bandyopadhyay, 2020. The atoxigenic biocontrol product Aflasafe SN01 is a valuable tool to mitigate aflatoxin contamination of both maize and groundnut cultivated in Senegal. Plant Dis., 104: 510-520.
- Mupangwa, W., I. Nyagumbo, F. Liben, L. Chipindu, P. Craufurd and S. Mkuhlani, 2021. Maize yields from rotation and intercropping systems with different legumes under conservation agriculture in contrasting agro-ecologies. Agric. Ecosyst. Environ., 306.
- Mabele, A.S., H.K. Were, M.F.O. Ndong'a and B. Mukoye, 2021. Occurrence and genetic diversity of groundnut rosette assistor virus in Western Kenya. Crop Prot., 139.
- Lengwati, D.M., C. Mathews and F.D. Dakora, 2020. Rotation benefits from N2-fixing grain legumes to cereals: From increases in seed yield and quality to greater household cash-income by a following maize crop. Front. Sustainability Food Syst., 4.
- Shalim Uddin, M., M.J. Rahaman, S.A. Bagum, M.J. Uddin and M.M. Rahaman, 2003. Performance of intercropping of maize with groundnut in saline area under rainfed condition. Pak. J. Biol. Sci., 6: 92-94.
- Nyaga, J., C.W. Muthuri, E. Barrios, I. Öborn and F.L. Sinclair, 2019. Enhancing maize productivity in agroforestry systems through managing competition: Lessons from smallholders’ farms, Rift Valley, Kenya. Agrofor. Syst., 93: 715-730.
- Srivastava, A.K., C.M. Mboh, B. Faye, T. Gaiser, A. Kuhn, E. Ermias and F. Ewert, 2019. Options for sustainable intensification of maize production in Ethiopia. Sustainability, 11.
- Reddy, K.R.N., B. Salleh, B. Saad, H.K. Abbas, C.A. Abel and W.T. Shier, 2010. An overview of mycotoxin contamination in foods and its implications for human health. Toxin Rev., 29: 3-26.
- Iqbal, S.Z., 2021. Mycotoxins in food, recent development in food analysis and future challenges; A review. Curr. Opin. Food Sci., 42: 237-247.
- Mahmood, Z.A., N. Shaheen, F. Tasleem, S. Imam, S. Abidi and I. Azhar, 2017. Detection of aflatoxins and use of scanning electron microscope for the identification of fungal species in some commonly used spices. Asian J. Plant Sci. Res., 7: 64-73.
- Negash, D., 2018. A review of aflatoxin: Occurrence, prevention, and gaps in both food and feed safety. J. Nutr. Health Food Eng., 8: 190-197.
- Pócsi, I., F. Giacometti, Á. Ambrus and A.F. Logrieco, 2020. Editorial: Aspergillus-derived mycotoxins in the feed and food chain. Front. Microbiol. 11.
- Bbosa, G.S., D. Kitya, A. Lubega, J. Ogwal-Okeng, W.W. Anokbonggo and D.B. Kyegombe, 2013. Review of the Biological and Health Effects of Aflatoxins on Body Organs and Body Systems. In: Aflatoxins-Recent Advances and Future Prospects, Razzaghi-Abyaneh, M. (Ed.), IntechOpen, United Kingdom, ISBN: 978-953-51-0904-4, .
- Cotty, P.J. and R. Jaime-Garcia, 2007. Influences of climate on aflatoxin producing fungi and aflatoxin contamination. Int. J. Food Microbiol., 119: 109-115.
- Bauchet, J., S. Prieto and J. Ricker-Gilbert, 2021. Improved drying and storage practices that reduce aflatoxins in stored maize: Experimental evidence from smallholders in Senegal. Am. J. Agric. Econ., 103: 296-316.
- Mahuku, G., H.S. Nzioki, C. Mutegi, F. Kanampiu, C. Narrod and D. Makumbi, 2019. Pre-harvest management is a critical practice for minimizing aflatoxin contamination of maize. Food Control, 96: 219-226.
- Were, E.O., E.N. Nyaboga and E.M. Ateka, 2019. Survey of the incidence and distribution of groundnut rosette disease in major groundnut-producing regions of Western Kenya. Afr. J. Biotechnol., 18: 1105-1112.
- Lewis, J.E., J. Rowland and A. Nadeau, 1998. Estimating maize production in Kenya using NDVI: Some statistical considerations. Int. J. Remote Sens., 19: 2609-2617.
- Nelson, M.C., M.W. Margaret and K.M. Lucy, 2016. Oil contents and aflatoxin levels in peanuts varieties produced in Busia and Kisii Central Districts, Kenya. Trop. Med. Surg., 4.
- Atsiaya, G.O., E.O. Gido and K.W. Sibiko, 2023. Uptake of climate-smart agricultural practices among smallholder sorghum farmers in Busia County, Kenya. Cogent Food Agric., 9.
- Maina, F.N.W., L.S. Gohole and R.M. Muasya, 2017. Effects of storage methods and seasons on seed quality of jute mallow morphotypes (Corchorus olitorius) in Siaya and Kakamega Counties, Kenya. Afr. J. Food Agric. Nutr. Dev., 17: 12395-12412.
- Ogenga, J.O., E.M. Mugalavai and N.O. Nyandiko, 2018. Impact of rainfall variability on food production under rainfed agriculture in Homa Bay County, Kenya. Int. J. Sci. Res. Publ., 8: 857-870.
- Olela, S.O., G.L. Makokha and K. Obiero, 2022. Spatiotemporal variability of temperature, relative humidity and rainfall in different altitudes of Lower Lake Victoria Basin, Kenya. J. Clim. Policy, 1: 1-19.
- Taherdoost, H., 2016. Sampling methods in research methodology; how to choose a sampling technique for research. Int. J. Acad. Res. Manage., 5: 18-27.
- Marete, G.N., L.W. Kanja, J.M. Mbaria, M.O. Okumu, P.A. Ateku, H. Korhonen and V. Joutsjoki, 2020. Effects of the use of good agricultural practices on aflatoxin levels in maize grown in Nandi County, Kenya. Sci, 2.
- Salano, E.N., M.A. Obonyo, F.J. Toroitich, B.O. Odhiambo and B.O. Aman, 2016. Diversity of putatively toxigenic Aspergillus species in maize and soil samples in an aflatoxicosis hotspot in Eastern Kenya. Afr. J. Microbiol. Res., 10: 172-184.
- Owino, A.A., J.O. Ochuodho and J.O. Were, 2013. Morphological diversity of net blotch fungi (Pyrenophora teres) infecting barley (Hordeum vulgare) in barley growing areas of Kenya. J. Exp. Biol. Agric. Sci., 1: 473-479.
- Were, J.O., P.C. Ngeywo, J.O. Ochuodho, N.K. Rop and A.A. Owino, 2015. Diversity and selectivity of mycotoxin fungi affecting Arachis hypogaea seed quality in Western Kenya. Int. J. Curr. Microbiol. Appl. Sci., 4: 216-221.
- Tranchida, M.C., N.D. Centeno and M.N. Cabello, 2014. Soil fungi: Their potential use as a forensic tool. J. Forensic Sci., 59: 785-789.
- Okayo, R.O., D.O. Andika, M.M. Dida, G.O. K’Otuto and B.M. Gichimu, 2020. Morphological and molecular characterization of toxigenic Aspergillus flavus from groundnut kernels in Kenya. Int. J. Microbiol., 2020.
- Manawasinghe, I.S., A.J.L. Phillips, J. Xu, A. Balasuriya and K.D. Hyde et al., 2021. Defining a species in fungal plant pathology: Beyond the species level. Fungal Diversity, 109: 267-282.
- Jayawardena, R.S., K.D. Hyde, A.R.G. de Farias, C.S. Bhunjun and H.S. Ferdinandez et al., 2021. What is a species in fungal plant pathogens? Fungal Diversity, 109: 239-266.
- Makhlouf, J., A. Carvajal-Campos, A. Querin, S. Tadrist and O. Puel et al., 2019. Morphologic, molecular and metabolic characterization of Aspergillus section Flavi in spices marketed in Lebanon. Sci. Rep., 9.
- Moslem, M.A., A. Mashraqi, K.A. Abd-Elsalam, A.H. Bahkali and M.A. El-Naggar, 2010. Molecular detection of ochratoxigenic Aspergillus species isolated from coffee beans in Saudi Arabia. Genet. Mol. Res., 9: 2292-2299.
- Saif, F.A., S.A. Yaseen, A.S. Alameen, S.B. Mane and P.B. Undre, 2020. Identification of Penicillium species of fruits using morphology and spectroscopic methods. J. Phys.: Conf. Ser., 1644.
- Upendra, R.S., P. Khandelwal, Z.R. Amiri, L. Shwetha and M.S. Ausim, 2013. Screening and molecular characterization of natural fungal isolates producing lovastatin. J. Microb. Biochem. Technol., 5: 25-30.
- Guerrero, J.J.G., T.U. Dalisay, I.B. Pangga and N.D. Organo, 2019. Siderophores of fungi from Lipa clay loam soil, Philippines. Philippine J. Syst. Biol., 13: 71-84.
- Ulloa, M., P. Lappe, S. Aguilar, H. Park, A. Pérez-Mejía, C. Toriello and M.L. Taylor, 2006. Contribution to the study of the mycobiota present in the natural habitats of Histoplasma capsulatum: An integrative study in Guerrero, Mexico. Rev. Mex. Biodivers., 77: 153-168.
- Virk, M.S., R. Ramzan, M.A. Virk, X. Yuan and F. Chen, 2020. Transfigured morphology and ameliorated production of six Monascus pigments by acetate species supplementation in Monascus ruber M7. Microorganisms, 8.
- Asan, A., 2003. Identification of common Aspergillus species. Turk. J. Bot., 27: 161-161.
- Wang, M., X.M. Tan, F. Liu and L. Cai, 2018. Eight new Arthrinium species from China. MycoKeys, 34: 1-24.
- Aghdam, S.A. and K.B. Fotouhifar, 2016. New reports of endophytic fungi associated with cherry (Prunus avium) and sour cherry (Prunus cerasus) trees in Iran. Mycologia Iran., 3: 75-85.
- Kebeish, R.M. and A.S. El-Sayed, 2012. Morphological and molecular characterization of L-methioninase producing Aspergillus species. Afr. J. Biotechnol., 11: 15280-15290.
- Jayshree, T. and J. Seema, 2018. Morphological and molecular study of Aspergillus tamarii Kita. Bull. Pure Appl. Sci. Bot., 37b: 124-129.
- Bandh, S.A., A.N. Kamili and B.A. Ganai, 2011. Identification of some Penicillium species by traditional approach of morphological observation and culture. Afr. J. Microbiol. Res., 5: 3493-3496.
- Al-Hmoud, N., M.A. Ibrahim, H. Al-Rousan and A. Alseyah, 2012. The prevalence of aflatoxinogenic Aspergillus parasiticus in Jordan. Int. J. Microbiol., 2012.
- Mahmoud, G.A.E. and A.N.A. Zohri, 2021. Amedment stable kojic acid produced by non-toxinogenic aspergillus oryzae using five levels central composite design of response surface methodology. J. Microb. Biotechnol. Food Sci., 10.
- Su, L., H. Deng and Y.C. Niu, 2016. Phialemoniopsis endophytica sp. nov., a new species of endophytic fungi from Luffa cylindrica in Henan, China. Mycol. Progress, 15.
- Lorenzo, J.M., P.E. Munekata, R. Dominguez, M. Pateiro, J.A. Saraiva and D. Franco, 2018. Main Groups of Microorganisms of Relevance for Food Safety and Stability: General Aspects and Overall Description. In: Innovative Technologies for Food Preservation: Inactivation of Spoilage and Pathogenic Microorganisms, Barba, F.J., A.S. Sant'Ana, V. Orlien and M. Koubaa (Eds.), Academic Press, United States, ISBN: 978-0-12-811031-7, pp: 53-107.
- Peterson, S.W., Ž. Jurjević and J.C. Frisvad, 2015. Expanding the species and chemical diversity of Penicillium section Cinnamopurpurea. PLoS ONE, 10.
- Teixeira, L.M., L. Coelho and N.D. Tebaldi, 2017. Characterization of Fusarium oxysporum isolates and resistance of passion fruit genotypes to fusariosis. Rev. Bras. Frutic., 39.
- Husain, A., Z. Hassan, N. Huda-Faujan, and M.N. Lani, 2017. Antifungal activity of lactic acid bacteria isolated from soil rhizosphere on Fusarium species infected chilli seeds. Am. Sci. Res. J. Eng. Technol. Sci., 29: 182-202.
- Kolařík, M., D.J. Spakowicz, R. Gazis, J. Shaw and A. Kubátová et al., 2017. Biatriospora (Ascomycota: Pleosporales) is an ecologically diverse genus including facultative marine fungi and endophytes with biotechnological potential. Plant Syst. Evol., 303: 35-50.
- Azeez, L.A., S. Muid and B.M. Hasnul, 2016. Identification of volatile secondary metabolites from an endophytic microfungus Aspergillus nomius KUB105. Malays. J. Anal. Sci., 20: 751-759.
- Majaa, I., M. Draganaa, I. Žarkob, K. Majac, V. Slobodana, N. Zoricaa and G.V. Jelicaa, 2018. Morphological and pathogenic properties of Fusarium proliferatum isolates: The causal agent of garlic (Allium sativum L.): Rot in Serbia. Ratarstvo Povrtarstvo, 55: 125-129.
- Xia, M.C., P. Bao, A.J. Liu, S.S. Zhang and T.J. Peng et al., 2018. Isolation and identification of Penicillium chrysogenum strain Y5 and its copper extraction characterization from waste printed circuit boards. J. Biosci. Bioeng., 126: 78-87.
- Rachniyom, H. and T. Jaenaksorn, 2008. Effect of soluble silicon and Trichoderma harzianum on the in vitro growth of Pythium aphanidermatum. J. Agric. Technol., 4: 57-71.
- Martínez-Culebras, P.V., M. Gandía, A. Boronat, J.F. Marcos and P. Manzanares, 2021. Differential susceptibility of mycotoxin-producing fungi to distinct antifungal proteins (AFPs). Food Microbiol., 97.
- Ropars, J., G. Aguileta, D.M. de Vienne and T. Giraud, 2014. Massive gene swamping among cheese-making Penicillium fungi. Microbial Cell, 1: 107-109.
- Gupta, C., S. Yadav, M. Sircar, R. Gupta and N. Chavhan et al., 2022. P204 secondary organizing pneumonia caused by Aspergillus flavus in immunocompromised patients. Med. Mycol., 60.
- Brauer, V.S., A.M. Pessoni, T.A. Bitencourt, R.G. de Paula, L. de Oliveira Rocha, G.H. Goldman and F. Almeida, 2020. Extracellular vesicles from Aspergillus flavus induce M1 polarizatio in vitro. mSphere, 5.
- Njoroge, S.M.C., 2018. A critical review of aflatoxin contamination of peanuts in Malawi and Zambia: The past, present, and future. Plant Dis., 102: 2394-2406.
- Monda, E., J. Masanga and A. Alakonya, 2020. Variation in occurrence and aflatoxigenicity of Aspergillus flavus from two climatically varied regions in Kenya. Toxins, 12.
- Houbraken, J., S. Kocsubé, C.M. Visagie, N. Yilmaz and X.C. Wang et al., 2020. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol., 95: 5-169.
- Houbraken, J., R.P. de Vries and R.A. Samson, 2014. Modern Taxonomy of Biotechnologically Important Aspergillus and Penicillium Species. In: Advances in Applied Microbiology, Sariaslani, S. and G.M. Gadd (Eds.), Academic Press, Cambridge, Massachusetts, ISBN: 9780128002629, pp: 199-249.
- Dorn, B., H.R. Forrer, E. Jenny, F.E. Wettstein, T.D. Bucheli and S. Vogelgsang, 2011. Fusarium species complex and mycotoxins in grain maize from maize hybrid trials and from grower’s fields. J. Appl. Microbiol., 111: 693-706.
- Casquete, R., A. Rodríguez, A. Hernández, A. Martín, T. Bartolomé, J.J. Córdoba and M.G. Córdoba, 2017. Occurrence of toxigenic fungi and mycotoxins during smoked paprika production. J. Food Prot., 80: 2068-2077.
- Mbaawuaga, E.M., J.O. Nwabude and D. Shiriki, 2020. Effects of farmers’ practices on maize (Zea mays) contamination by potential aflatoxigenic fungi and aflatoxin in Benue State, Nigeria. Agric. Sci., 11: 500-513.
- Smith, L.E., M. Stasiewicz, R. Hestrin, L. Morales, S. Mutiga and R.J. Nelson, 2016. Examining environmental drivers of spatial variability in aflatoxin accumulation in Kenyan maize: Potential utility in risk prediction models. Afr. J. Food Agric. Nutr. Dev., 16: 11086-11105.
- Hawkins, L.K., G.L. Windham and W.P. Williams, 2005. Effect of different postharvest drying temperatures on Aspergillus flavus survival and aflatoxin content in five maize hybrids. J. Food Prot., 68: 1521-1524.
- Kaale, L.D., M.E. Kimanya, I.J. Macha and N. Mlalila, 2021. Aflatoxin contamination and recommendations to improve its control: A review. World Mycotoxin J., 14: 27-40.
- Meijer, N., G. Kleter, M. de Nijs, M.L. Rau, R. Derkx and H.J. van der Fels-Klerx, 2021. The aflatoxin situation in Africa: Systematic literature review. Compr. Rev. Food Sci. Food Saf., 20: 2286-2304.
- Stepman, F., 2018. Scaling-up the impact of aflatoxin research in Africa. The role of social sciences. Toxins, 10.
- Costa, J., R. Rodríguez, E. Garcia-Cela, A. Medina and N. Magan et al., 2019. Overview of fungi and mycotoxin contamination in Capsicum pepper and in its derivatives. Toxins, 11.
- Singh, R., G. Kaur and G. Kaur, 2022. Shelf-life prolongation of spring groundnut pods (Arachis hypogaea L.) using packaging systems. J. Sci. Ind. Res., 81: 393-401.
- Birgen, J.K., R.C. Cheruiyot and T.E. Akwa, 2020. Mycotoxin contamination of stored maize in Kenya and the associated fungi. J. Plant Pathol. Res., 2: 7-13.
- Matumba, L., S. Namaumbo, T. Ngoma, N. Meleke, M. de Boevre, A.F. Logrieco and S. de Saeger, 2021. Five keys to prevention and control of mycotoxins in grains: A proposal. Global Food Secur., 30.
- Niyibituronsa, M., C. Mukantwali, M. Nzamwita, G. Hagenimana and S. Niyoyita et al., 2020. Assessment of aflatoxin and fumonisin contamination levels in maize and mycotoxins awareness and risk factors in Rwanda. Afr. J. Food Agric. Nutr. Dev., 20: 16420-16446.
- Fouché, T., S. Claassens and M. Maboeta, 2020. Aflatoxins in the soil ecosystem: An overview of its occurrence, fate, effects and future perspectives. Mycotoxin Res., 36: 303-309.
How to Cite this paper?
APA-7 Style
Mwatabu,
E.M., Were,
J.O., Chiveu,
J.C., Ouma,
E.O., Obonyo,
M. (2024). Diversity and Environmental Specificity of Fungal Isolates from Soils and Cereal Grains in Western Kenya. Trends in Agricultural Sciences, 3(1), 38-51. https://doi.org/10.17311/tas.2024.38.51
ACS Style
Mwatabu,
E.M.; Were,
J.O.; Chiveu,
J.C.; Ouma,
E.O.; Obonyo,
M. Diversity and Environmental Specificity of Fungal Isolates from Soils and Cereal Grains in Western Kenya. Trends Agric. Sci 2024, 3, 38-51. https://doi.org/10.17311/tas.2024.38.51
AMA Style
Mwatabu
EM, Were
JO, Chiveu
JC, Ouma
EO, Obonyo
M. Diversity and Environmental Specificity of Fungal Isolates from Soils and Cereal Grains in Western Kenya. Trends in Agricultural Sciences. 2024; 3(1): 38-51. https://doi.org/10.17311/tas.2024.38.51
Chicago/Turabian Style
Mwatabu, Edward, Mwakichwa, Javan Omondi Were, Josiah Chemulanga Chiveu, Evans Ochieng Ouma, and Meshack Obonyo.
2024. "Diversity and Environmental Specificity of Fungal Isolates from Soils and Cereal Grains in Western Kenya" Trends in Agricultural Sciences 3, no. 1: 38-51. https://doi.org/10.17311/tas.2024.38.51

This work is licensed under a Creative Commons Attribution 4.0 International License.