Morphologicaand Phytochemical Characterization of Cassiinae in Nigeria
-
Okpara Onyinyechi

Department of Botany, Joseph Sarwuan Tarka University, Makurdi, Nigeria
Abiodun AyodeleUniversity of Ibadan, Oyo State, Nigeria
| Received 20 Jun, 2024 |
Accepted 16 Sep, 2024 |
Published 30 Sep, 2024 |
Background and Objective: Cassiinae is the largest subtribe that constitutes the tribe Cassieae (Caesalpiniodeae) in the Fabaceae family. The autonomy and classification of the generic categories (Cassia, Senna and Chamaecrista) in the subtribe were established in this work using multiple character variations obtained from anatomical, morphological and phytochemical evidence to determine the taxonomic relationship among 10 studied taxa. This has led to the production of a reliable identification key. Materials and Methods: A total of 45 vegetative and reproductive characters, 13 anatomical and 13 phytochemical characters were qualitatively and quantitatively evaluated. The characters obtained were subjected to numerical and principal component analysis following Sokal and Sneath principles. The phenetic relationship among the groups was determined by hierarchical cluster analysis using the R programme. Results: A dendrogram containing three groups and a few subgroups representing the 10 taxa was produced. Cluster 1 comprises Cassia fistula and Cassia sieberiana. Cluster 2 contains only Chamaecrista rotundifolia while cluster 3 includes Senna alata, Senna hirsuta, Senna occidentalis, Senna siamea, Senna polyphylla, Senna obtusifolia and Senna surattensis. The resultant delimitation into 3 groups is a result of the combination of OTU of gross morphological and phytochemical characters during numerical analysis. The two-dimensional model of principal component analysis used highlighted shared characters of taxonomic significance that support delimitation of members of the group. Conclusion: This study affirms that morphological features are not sufficient enough but indispensable in the identification and classification of a clade.
| Copyright © 2024 Onyinyechi and Ayodele. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Cassiinae is one of the five subtribes in tribe Cassieae (Carotoniinae, Dialiinae, Duparquetiinae, Cassiinae and Labicheinae) and is notably the largest in terms of number of species consisting of about 740 species. The subtribe is categorized into 3 genera namely: Cassia, Senna and Chamaecrista and belongs to the subfamily Caesalpinoideae in the family Fabaceae. They are mostly cultivated for their economic, medicinal and ecological benefits as they promote food security, sustainable agricultural practices, sustenance of biodiversity and climate change1. Members of the subtribe are rich in phytochemical compounds useful in traditional medicine and the pharmacological industry. They are usually recognized by their showy flowers; thus, they are aesthetically relevant2. The sub-tribe are distributed in both wet and dry regions with a few in the temperate regions of the world3. The systematic study of chemical variation in plant taxa is useful in the classification of plants based on their specific secondary metabolites and biosynthetics4. There are three broad categories of compounds used in the classification of plants namely; primary metabolites, secondary metabolites and semantides4. It has been proven that certain compounds and related substances may be characteristic of certain taxonomic groups and evidently, the pathway of chemical evolution can be established since chemical substances are retained by metabolic processes5. Compounds retained by groups or ancestors, may lead to the understanding of relationships within and between groups6,7. Information obtained from this method is best used in conjunction with other sources of taxonomic evidence such as morphology, anatomy, cytological and molecular data to establish a system of classification that reflects natural relationships as accurately as possible. Based on the critical comparative study carried out in this work variations and similarities range from the gross morphological, anatomy and phytochemical characters obtained to serve as a tool for finding distinctive diagnostic characters that complemented and enhanced the delimitation and classification of members of the group. Thus, the combination of gross morphological and phytochemical characters in this study provided taxonomic evidence useful to trace evolutionary relationships within the taxa using the natural system of classification.
This study aimed to examine the taxonomic relationships among selected species in the subtribe Cassiinae. The specific objectives of this study were to:
| • | Determine the gross morphological and phytochemical character variations among studied taxa | |
| • | Determine the phenetic relationships among taxa using gross morphological and phytochemical evidences | |
| • | Provide a good and reliable taxonomic key for the recognition of members of the subtribe Cassiinae |
MATERIALS AND METHODS
Plant collection: This study was carried out from May, 2021 to January, 2022. Ten species representing three genera in the sub-tribe Cassiinae (Fabaceae: Caesalpinoideae) in Nigeria were collected and determined into species based on the keys to species and classified into genus according to Irwin and Barneby8. Herbarium species housed at the University of Ibadan Herbarium and Forest Research Institute of Nigeria (FRIN) were used to confirm specimens by comparison with fresh specimens and used for morphometric studies. The specimen collected includes Chamaecrista rotundifolia (Pers.) Greene, Senna hirsuta (L.) H.S. Irwin & Barneby, Senna alata (L.) Roxb, Senna occidentalis (L.) Link, Senna siamea (Lam.) H.S. Irwin & Barneby, Cassia fistula L., Cassia sieberiana DC, Senna obtusifolia (L.) H.S. Irwin & Barneby, Senna polyphylla (Jacq.) H.S. Irwin & Barneby and Senna surattensis (Burm.f.) H.S. Irwin & Barneby.
Leaf morphometrics: Vegetative and floral macro-morphological characters of species in the Cassiinae subtribes were examined in this study. The quantitative characters determined include leaflet width and length, petiolule length, blade/rachis length ratio and leaf length/width ratio. The qualitative characteristics examined were the type of inflorescence and position, leaf arrangement, venation, shape, margin, surface beneath and above, texture, apex, base, fruit shape and color. The margin, length and width of specimens under study were measured using a twine and meter rule.
Anatomical studies
Preparation of leaf epidermal peels: Fresh leaves of each species were preserved in 50% ethanol and rinsed in ordinary water. About 1 cm2 cut from the standard median portion of the leaf of each specimen was made. Each specimen was put in a glass petri dish and concentrated nitric acid was added so that the leaf was immersed in acid. These were left in the sun outside the laboratory to hasten the action of the acid. The formation of air bubbles in the leaves indicated the separation of the upper and the lower epidermis from the mesophyll. The specimens were then transferred into a new Petri-dish and rinsed several times in distilled water. The epidermis for each specimen was separated with a pair of forceps and cleaned with a camel hair brush by removing the residual mesophyll layer9. Specimens were counter-stained with Alcian blue for another 5 min. Stained sections were mounted on microscope slides with 2 drops of 25% glycerol added and covered with a cover slip.
Preparation of petiolule section: Petiotule sections were made using a rotary microtome ‘820’. Sectioned specimens were cleared with 5% sodium hypochlorite to remove pigments and then rinsed in distilled water. The very thin sections were collected and stained in safranin for 5 min. The specimens were counter-stained with Alcian blue for another 5 min. Stained sections were mounted on clear microscope slides with 2 drops of 25% glycerol added and covered with a cover slip. The slides were previewed under the microscope (Olumpus corperation, Shinjuku Monolith, 3-1, Nishi Shinjuku 2-chrome, Shinjuku-Ku, Tokyo, Japan) and the edges were carefully sealed with transparent nail polish. Photomicrographs of the slides were captured using an Olympus CX 31 photomicroscope with a built-in digital camera10.
Phytochemical analysis: The qualitative and quantitative phytochemical characters were analyzed and determined using fresh leaves of the ten taxa identified in the herbarium were air-dried at a temperature between 36-38°C for two weeks. The dried samples were ground to powder and tested for the presence of phytochemical constituents following the following standard methods described as follows.
Alkaloid determination: Standard measurement procedures were used to weigh 5 g of fine powder of samples, weighed 250 mL of 10% acetic acid into ethanol and added to a covered beaker and allow to stand for 4 hrs. The filtered extract concentrates were made to pass through a water bath at 55°C to one-quarter of the original volume. The concentrated ammonium hydroxide was added to the extract in drops until the precipitation was complete. Then carefully collected and washed in dilute ammonium hydroxide and filtered. The residue obtained was dried and weighed. The weighed residue becomes the alkaloids11.
Determination of saponin content: Grounded samples of fine powder weighing 20 g were put into a conical flask and 100 cm3 of 20% aqueous ethanol was added and heated over a hot water bath at 55°C for 4 hrs with continuous stirring. The mixture was filtered and the residue was re-extracted with another 200 mL of 20% ethanol. The combined extracts were reduced to 40 mL over water bath at 90°C. The concentrate was then transferred into a 250 mL separatory funnel and 20 mL of diethyl ether was added and continuously shaken. The aqueous layer was retained while the ether layer was disposed of. The purification was repeated. This time about 60 mL of n-butanol was added. The combined n-butanol extracts were washed twice with 10 mL of 5% aqueous sodium chloride. The retained solution was heated in a water bath. After evaporation, the samples were dried in an oven to a constant weight. The dried samples are the saponins and is calculated as percentage12.
Determination of total phenolic content: The concentration of phenolic content in dried samples of Cassiinae extracts was evaluated using standard spectrophotometric method of Folin-Ciocalteu assay. The reaction mixture consisting of 1 mL of extract and 9 mL of distilled water was taken into a volumetric flask measured at 25 mL, 1 mL of Folin-Ciocalteu phenol reagent was treated to the mixture. The volume was set at 25 mL. Standard solutions of gallic acid measuring (20, 40, 60, 80 and 100 μg/mL) were prepared and incubated for 90 min at room temperature and the absorbance for tests and standard solutions determine against the reagent blank at 550 nm with Infitek UV-visible. spectrophotometer SP-MUV6000 (Jinan, Shandong, China). The total phenolic content was expressed as mg of GAE/g of extract.
Determination of tanin content: The tannins were determined following standard methods by Folin-Ciocalteu. About 0.1 mL of the grounded plant extract was added to a volumetric flask (10 mL) containing 7.5 mL of distilled water and 0.5 mL of Folin-Ciocalteu, phenol reagent, 1 mL of 35% Na2CO3 solution and diluted to 10 mL with distilled water. The mixture was shaken vigorously and kept at room temperature of about 37-38°C for 30 min. Standard solutions of gallic acid (20, 40, 60, 80 and 100 μg/mL) were prepared in the same manner as previous procedures explained earlier. The absorbance test and standard solutions weres measured against the blank at 75 nm with Infitek UV-visible spectrophotometer SP-MUV6000 (Jinan, Shandong, China). The tannin content measurements were expressed in terms of mg of GAE/g of extract13.
Determination of total flavonoid content: The total flavonoid content was determined using the aluminium chloride colorimeter assay. The reaction mixture is made up of 1 mL of extract and 4 mL of distilled water was taken in a 10 mL volumetric flask. In the flask, 0.30 mL of 5% sodium nitrite was treated and after 5 min, 0.3 mL of 10% aluminium chloride was mixed. After 5 min, 2 mL of 1 M sodium hydroxide was treated and diluted to 10 mL with distilled water. Standard solutions of quercetin (20, 40, 60, 80 and 100 μg/mL) were prepared. The absorbance for test and standard solutions was determined against the reagent blank at 510 nm with Infitek UV-visible spectrophotometer SP-MUV6000 (Jinan, Shandong, China). The total flavonoid content was expressed as mg of QE/g of extract14.
Determination of glycoside content: The total glycoside content was determined by adding 2.00 g of grounded sample to 20 cm3 of water and heated for 5 min in a water bath, then, filtered using the Gem filter paper (12.5 cm). The following tests were carried out on the filtrate:
| • | Add 0.2 cm3 of Fehling’s solutions A and B were mixed with (5 cm3) of the filtrate until it became alkaline (tested with litmus paper). A brick-red coloration upon heating showed presence of glycoside | |
| • | About 15 cm3 of 1.0 M sulphuric acid was used to repeat the test in (a) and the variation in quantity of precipitate was evaluated and compared with that of test (a). High precipitate content in test B indicates the presence of glycoside15 |
Determination of terpenoid content: Measurement of the terpenoid content was investigated using the analytical method. Each powdered sample about (0.30 g) was weighed into a beaker and extracted for 2 hrs. A mixture of chloroform (2 cm3) and concentrated tetra-oxo sulphate (VI) acid (3 cm3) was added to 5 cm3 of each extract to form a layer. The presence of a reddish-brown coloration at the interface indicates positive results for the presence of terpenoids16.
Determination of steroids and phyto-steroids content: Glycoside is dissolved in 1% ferric sulphate solution in (5%) glacial acid. One or two drops of concentrated sulphuric acid are added. A blue colour develops due to the presence of deoxy sugar17.
Determination of cardiac glycosides (Keller Killiani test): To 1 mL of plant extract equal volume of chloroform is added and subjected to 2-3 drops of concentrated sulphuric acid. The appearance of a brown ring indicates the presence of steroids and the appearance of bluish brown ring indicates the presence phyto-steroid17.
Determination of anthraquinone: To determine the presence of anthraquinone content about 5.0 g of each sample extract were shaken with 10 mL of benzene in a test tube and filtered. As 5 mL of 10% ammonia was added to the filtrate. The mixture was shaken vigorously to obtain. pink, red or violet colour in the lower portion of the ammonia. This is an indication of the presence of free hydroxyl-anthraquinone17.
Determination of quinones content: To 1 mL of plant extract, 1 mL of concentrated sulphuric acid was added. The formation of red color indicates the presence of quinones17.
Determination of coumarin content: To 1 mL of plant extract, 1 mL of 10% NaOH was added. The formation of yellow colour indicates the presence of coumarins13.
Data analysis: The data generated from the morphological and phytochemical characters were analyzed using descriptive statistics. Hierarchical cluster and principal component analysis were carried out to show the phenetic relationships among the taxa. Pair group clustering was measured using the R software version 4.4.0 for statistical computing and the graphics13.
Principal component analysis (PCA): Principal component analysis (PCA), was used to examine relationships among several quantitative variables:
where, Y is matrix comprising of 10 species from Cassiinae and 70 characters, X is matrix scores and B is matrix of Eigen vectors (latent vectors) which are themselves calculated from the data matrix of the character scores. The minimum Eigen criterion was used to select how many factors can best explain the matrix18.
RESULTS
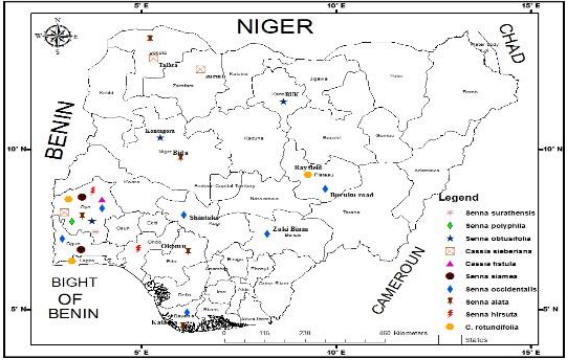
Macro-morphological studies: Vegetal and floral morphological characteristics were examined in ten species belonging to the subtribe Cassinae in Nigeria. The locality, collectors’ name, voucher number and date of collection of specimens were inventoried from two herbaria in Ibadan, Oyo State (Forest Institute of Nigeria and University of Herbarium) as shown in Table 1. Results from Table 1, were used to produce a species distribution map illustrated in Fig. 1. All species investigated were found dominant in the Southwestern Region of Nigeria. Senna obtusifolia is dominant in the North Eastern region. While Senna alata, Senna obtusifolia and Cassia sieberiana are prominent in the North Western part. Chamaecrista rotundifolia and Senna occidentalis are the species commonly distributed in the North Central Guinea Savanna Region. Figure 2a-j were showing photograph of Cassiinae species in their natural habitat. Table 2 shows the localities of operational taxonomic units (OTUs) used for micro-morphological and phytochemical studies were all within the Ibadan metropolis as inventoried. Table 3 shows the qualitative macro-morphological characteristic features determined for the studied specimen. A total of 38 qualitative characters involving both vegetal and floral characters were examined. Varying features such as habit type, leaflet arrangement, venation, presence or absence of stipule, leaflet shape margin, apex and base, fruit color and seed shape, seeds per lobe number and inflorescence position were evaluated. Leaves of members of Cassiinae are characteristically alternately arranged, venation pinnate with presence or absence of stipule. Leaflet shapes are asymmetrical, sub-rotund or broadly obovate. Leaflet margins are majorly entire with apex round, acute or acuminate. The leaflet base is mostly rounded or cuneate. The fruit and seed color are brown at maturity, with several seeds per lobe ranging from 20-100 in number. The inflorescence is mostly racemose and scarcely paniculate, positioned either as terminal or axillary. The seed shapes of species vary as either flattened, obovate, or rhomboid. Table 4 shows results from measurements on the quantitative macro-morphological characters considered. Several observed variations such as leaflet length and width ranging from 0.62-29.62 and 0.34-9.86 cm, rachis (2.40-38.50 cm), petiolule (0.10-0.50 cm) and blade lengths (1.54-51.48 cm) were evaluated in each species and their mean values and standard error recorded.
|
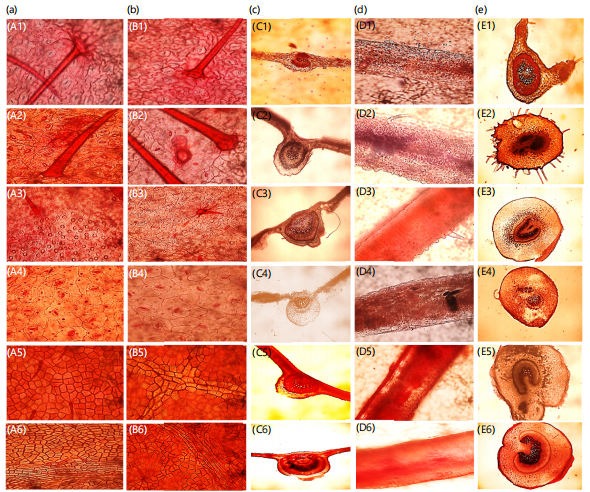
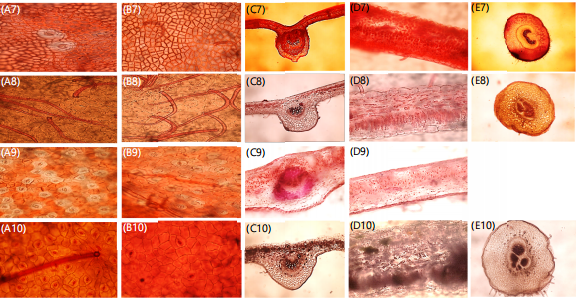
Anatomical studies: Figure 3a-e show the photomicrograph of epidermal leaf surfaces (adaxial and abaxial), main vein, lamina and petiolule of studied specimens. Figure 3 showed a summary of all the micro-morphological features examined for each OTUs studied. Table 5 describes the qualitative micro-morphological characters examined. Descriptive features such as cell shape, stomata shape, anticlinal wall pattern, cell wall striation, presence or absence of trichomes and presence or absence of crystals were investigated. The stomatal cells of examined specimens were mostly anisocytic and anomocytic. Cell shapes varied as either irregular or polygonal. Trichomes varied on either adaxial and abaxial surfaces of the epidermal cells, interestingly, trichomes were absent in Cassia fistula, Cassia sieberiana and Senna polyphylla. Crystalic structures were observed in Senna siamea, Cassia fistula and Senna surattensis. Table 6 shows the quantitative micro-morphological characters evaluated. The highest stomata index value was recorded in Senna alata (91.9%) and lowest in Senna surattensis (7.7%). The stomata density ranged from (0.00-102.5 m/sec2). The highest stomatal length was evaluated in Chamaecrista rotundifolia (10.63 μm) and the lowest in Senna obtusifolia (0.75 μm). Variations in the transverse section of the lamina showed double-layer thick cells in all species examined except in Chamaecrista rotundifolia. Cell shapes in studied species were mostly cuboidal except in C. rotundifolia which had an elongated type. The outlines in the main vein were mostly protruding and shaped and were either elongated or cuboidal in most cases. Epidermis in the main vein was mainly collateral in all with exception of Senna alata which differed as bicollateral. Parenchymatous cortex in the main vein ranges from crescent to heart-shape with unique circular types found only in Senna hirsuta. The micro characters of the petiolule examined in all specimen indicated varying outline shapes such as U-shaped in OTU 1, 2, 3,4 5, 6, 7, 8, 9 and 10, undulated outline in S. hirsuta and C. Sieberiana and flat outline surfaces in S. siamea, S. obtusifolia and S. surattensis. Also, the sclerenchyma cells in the petiolule were mostly thin and scarcely thick with majority having small pith except in Chamaecrista rotundifolia. Delimitation within species using the petiolule characters were not holistic because of the absence of a well-defined petiolule structure in Senna polyphylla.
 
|
|
| Table 1: | List of herbarium specimens used for macro-morphological studies | |||
| Taxa | Locality | Collector’s name | Collector(s)/voucher number | Date |
| Chamaecrista rotundifolia (Pers.) Greene | Bridge Embarkment Waterworks Ogbomosho Oyo | Dr. J. Lowe 799 | UCI 2019 | 17/12/1965 |
| University of Ibadan Campus | Dr. J. Lowe 1017 | UIH 2020 | 14/10/1966 | |
| Roadside, Appleton Road, University of Ibadan | P.W. Richards | UIH 19181 | 03/11/1980 | |
| Jos | G.S. Puri | UIH 2014 | 14/10/1966 | |
| Between Olobeweji and Ado Awaye | P.W. Richards | UIH 2013 | 03/11/1980 | |
| Oyo Iseyin-Ado Road | A.A. Lady | UIH 2012 | 31/10/1962 | |
| Ilora Oyo | G.W. Iveus | UIH 13316 | 12/04/1972 | |
| Rayfield Jos Division | W.J. Howard | UIH 16448 | 19/08/1973 | |
| Benue-Plateau State | Dr. G.S Puri. | UIH 2015 | 01/12/1962 | |
| Gambari Forest Reserve | Sharma | UIH 19099 | 12/08/1971 | |
| Lagos Waste Ground | Dr. J. Lowe 485 | UIH 2018 | 24/11/1964 | |
| Senna hirsuta (L.) H.S. Irwin & Barneby | 17 Miles Ore-Benin Province, Ondo District | J.F. Barker | UIH 1988 | 17/09/1965 |
| Barthroad Student Village, UI | L.O. Ogunniran | UIH 20111 | 10/01/1983 | |
| UI Second Gate by Road Side | A.O. Chivinge | UIH 19905 | 13/10/1978 | |
| Student Village UI | K.T. Kumi | UIH 20106 | 10/01/1983 | |
| UI Campus near Fish Pond | Aghachi | UIH 18159 | 06/01/1976 | |
| Behind Botany Nursery | E.J. Enamuotor | UIH 18176 | 28/01/1974 | |
| Behind Zoological Garden | A.M. Salaam | UIH 19919 | 13/10/1978 | |
| Senna alata (L.) Roxb. | Adeagba A.T. Apete Ibadan | E.A. Olayinola | UIH 23005 | 11/05/2021 |
| Okomu Forest National Park | M.K. Ayodele | UIH 22671 | 04/04/2018 | |
| Sokoto | E. Willie | UIH 14265 | 29/10/1971 | |
| Botany Department, UI | O.B, Oluwatobi | UIH 22376 | 13/10/2013 | |
| Department of Botany | Allsworth Jones and Rosen | UIH 20072 | 13/02/1983 | |
| Kaiama Town, Kolokumi Area, Yenagoa Division | K.R.M. Williamson | UIH 15344 | 27/10/1973 | |
| Pici Village 8 km West of Bida | R.M. Blench | UIH 19635 | 28/01/1982 | |
| Botany Department | A.A. Ogunshe | UIH 22291 | 2008 | |
| Senna occidentalis (L.) Link | Ibadan | O.O. Omotayo | UIH 22503 | 22/03/2016 |
| University of Ibadan | C.I. Ogbonna | UIH 20144 | 1983 | |
| Ibadan | K. Williamson | UIH 12968 | 17/06/1971 | |
| Shintako Kabba | J.F. Redhead | UIH 21008 | 21/08/1964 | |
| Near Kaiama in Town, Kolokuma Area (Yenagoa) | K.R.M. Williamson | UIH 15342 | 27/10/1973 | |
| Ijebu Ode | J.A. Okusi | UIH 2005 | 28/08/1935 | |
| Eleiyele Ibadan | Bamiyo | UIH 13433 | 10/07/1963 | |
| Zaki, Biam, Benue Province | D. Vermeer | UIH 16731 | Oct-75 | |
| Jos Bukuru Road | Dr. G.S. Puri | UIH 2007 | 25/07/1963 | |
| Senna siamea (Lam.) H.S. Irwin & Barneby | Ejioku, North East Ibadan | A.B. Oyelami | UIH 21700 | 08/12/1991 |
| University of Ibadan Campus | J. Lowe | UIH 20053 | 02/02/1983 | |
| Ijebu Ode | J.A. Okusi | UIH 2021 | 30/08/1937 | |
| Cassia fistula L. | BOT/MIC Department UI | J.B. Macjob | UIH 21615 | 23/05/1991 |
| Botany Department UI | Ezomoe Evelyn | UIH 21587 | 02/05/1991 | |
| Cassia sieberiana DC. | Behind Trenchard Hall, University of Ibadan | J. Lowe | UIH 20325 | 29/02/1984 |
| Aworo F.R. in High Forest in Family Open Place on Well Drained | B.O. Davimica | FHI 0027923 | 24/10/1968 | |
| Gradualslope, Growing among Dialium celtis and Antiaris | ||||
| LAUTECH Premises Open Space | R.W.J. Keay and Forest Students | FHI 0109967 | 24/04/2014 | |
| Zurmi Katsina Motor Road about 2 Miles from Zurmi | of Ibadan Forest school | FHI 0016135 | 12/04/1946 | |
| Talata Mafora by Zaria-Sokoto Road | R.W.J. Keay | FHI 007555-2 | 26/06/1944 | |
| Forest Hill, in Rest House Compound | J. Olorunfemi | FHI 0031502 | 13/101/1958 | |
| Arboretum Forest Research Ibadan | H. Donyeachum | FHI 0054745 | 07/01/1965 | |
| Forest Research Headquarters’ compound, Ibadan. | M.G. Latilo | FHI 043899-1 | 15/03/1960 | |
| Senna obtusifolia (L.) H.S, Irwin & Barneby | Zaria Railway Side | G. Jochson | UIH 14264 | 30/11/1972 |
| Near Pond on New Site Bayero University, Kano | J. Lowe | UIH 20470 | 02/12/1984 | |
| Salka, West of Kontagora. | ||||
| Back of Physics | R.M. Blench | UIH 19595 | Nov-81 | |
| Department, UI | P.P. Adeonipekim | UIH 21713 | 14/12/1991 | |
| Kusoji Mokwa-Kontagora Road | Sharma | UIH 20978 | 19/09/1977 | |
| Zaria-Kano Road | Sharma | UIH 20952 | 04/09/1977 | |
| Senna polyphylla (Jacq.) H.S. Irwin & Barneby | In front of main library, Botany University of Ibadan | J.O. Inegbedion | UIH 6630 | 09/11/1977 |
| Garden of British Deputy High Commissioner, Bodija, Ibadan | J. Lowe | UIH 18573 | 10/04/1979 | |
| Akwanga Keffi Road | S.O. Oyewole | UIH 19050 | Nov-75 | |
| Senna surathensis (Brum. F.). Irwin & Barneby | Gambari Forest Reserve, Teak Plantation Area | Mr A. Adebambo | FHI 0108182 | 14/12/2007 |
| University of Ibadan, Botanical Gardens | M.G latilo | FHI 0043497 | 23/10/1959 | |
| Ibadan University Medical College (U.M.C) Compound Tree | R.W.J. Keay | FHI 0025685 | 14/03/1950 |
| Table 2: | Operational taxonomic units (OTUs) and locality used for morphological and phytochemical studies | |||
| OTUs | Species | Locality |
| 1 | Chamaecrista rotundifolia (Pers.) Greene | Behind Botanical Garden, University of Ibadan, Ibadan |
| 2 | Senna hirsuta (L.) H.S. Irwin & Barneby | Agric Farm, University of Ibadan |
| 3 | Senna alata (L.) Roxb. | Abadina, shoe maker junction and front of Zik Hall, both inside University of Ibadan Campus |
| 4 | Senna occidentalis (L.) Link | Barika, Agbowo, Opposite University of Ibadan second gate |
| 5 | Senna siamea (Lam.) H.S. Irwin & Barneby | Besides Agric Farm University of Ibadan |
| 6 | Cassia fistula L. | Department of Botany, University of Ibadan |
| 7 | Cassia sieberiana DC. | Department of Agronomy, University of Ibadan |
| 8 | Senna obtusifolia (L.) H.S. Irwin & Barneby | Bank of Ajibode River, Behind Botanical Garden, University of Ibadan |
| 9 | Senna polyphylla (Jacq.) H.S. Irwin & Barneby | Department of Botany, University of Ibadan |
| 10 | Senna surattensis (Burm. F.). Irwin & Barneby | Botanical Garden, University of Ibadan |
| Table 3: | Qualitative macro-morphological characters of species in Cassiinae | |||
| Character | Chamarcrista rotundifolia | Senna hirsuta | Senna alata | Senna occidentalis | Senna siamea | Cassia fistula | Cassia sieberiana | Senna obtusifolia | Senna polyphylla | Senna surattensis |
| Habit Leaflet shape |
Herbaceous Asymmetrically sub-rotund to broad obovate |
Herb Lanceolate |
Shrub Oblong-obovate |
Herb Ovate to ovate-oblong |
Tree Oblong to ovate-oblong |
Tree Ovate |
Tree Elliptic-oblong |
Herb/Undershrub Obovate |
Shrub Elliptic-oblong |
Shrub Ovate to ovate-oblong |
| Leaflet margin | Entire | Entire | Entire | Entire | Entire | Entire | Entire | Entire | Entire | Entire |
| Leaflet apex | Round/mucronate | Acutely acuminate |
Rounded | Aute or acuminate | Rounded | Acute | Subacute | Acute | Rounded | Bluntly emarginate |
| Leaflet base | Asymmetrical | Acute or round | Obliquely truncated or semi-cordate |
Obtuse | Rounded to cuneate |
Cuneate | Rounded or broadly cuneate |
Cuneate to acute | Rounded to cuneate |
Cuneate to rounded |
| Leaflet surface above |
Ciliate | Villous | Glabrous | Glabrous | Glabrous | Glabrous and shining |
Paler and rarely pubescent |
Glabrous | Glabrous | Glabrous |
| Leaflet surface beneath |
Pubescent | Villous | Carinate | Dull glabrous | Rough to delicately pubescent |
Glabrous | Glabrous | Pubscent | Glabrous | Appressed pubscent |
| Leaflet texture | Membranous | Chartaceous | Chartaceous | Membranous | Sub coriaceous | Sub coriaceous | Sub coriaceous | Membranous | Chartaceous to membranous |
Chartaceous to membranous |
| Leaflet venation |
Pinnate | Pinnate | Pinnate | Pinnate | Pinnate | Pinnate | Pinnate | Pinnate | Pinnate | Pinnate |
| Leaflet arrangement |
Alternate | Alternate | Alternate | Alternate | Alternate | Alternate | Alternate | Alternate | Alternate | Alternate |
| Leaf forms | Pinnately compound | Pari pinnately compound |
Pari pinnately compound |
Pari pinnately compound |
Pari pinnately compound |
Pari pinnately compound |
Pari pinnately compound |
Pari pinnately compound |
Even pinnately compound |
Pari pinnately compound |
| Petiole | Round | Stout | Terete | Subterete | Slightly grooved | Round | Elliptic,ovate, obovate-elliptic |
Round | Pubscent | Sturdy |
| Petiole surface | N/A | Villous | Pubscent | Glabrous | Glabrous | Glabrous | Finely pubescent | Glabrous | Pubscent | Glabrous |
| Stipule shape | Lanceolate -cordate | Triangular | Asymmetrically triangular |
Lanceolate | Subulate | Deltoid | Narrowly triangular | Linear | Subulate | Linear falcate |
| Stipule surface | Fallacious | Densely pubescent |
Glabrous | Glabrous | Pubscent | Glabrous | Caducous | Membranous | Glabrous | Pubscent |
| Floral gland | Present | Present | Present | Present | Absent | Absent | Absent | Present | Present | Present |
| Bract | Persistent | Dischitous | Obovate | Lanceolate | Obovate | Ovate | Linear lanceolate | Linear acute | Ovate acute | Ovate acute |
| Bracteole | Present | Present | Absent | Absent | Absent | Present | Present | Absent | Absent | Present |
| Sepal shape | Lanceolate | Obovate/rounded | Obovate | Ovate | Rounded-ovate | Ovate, blunt | Elliptical | Ovate | Ovate | Ovate to orbicular |
| Sepal surface | Ciliate | Membranous- glabrous |
Membranous | Membranous | Pubscent | Glabrous | Slightly pubescent | Membranous | Membranous | Glabrous |
| Petal shape | Obovate | Obovate | Obovate to orbicular |
Obovate to spathulate |
Orbicular to obovate |
Obovate | Elliptical or ovate | Ovate, obtuse or round |
Ovate | Ovate to oblong |
| Petal surface | Glabrous | Membranous to glabrous |
Glabrous | Glabrous | Glabrous | Glabrous | Glabrous | Glabrous | Glabrous to pubescent |
Pubscent |
| Character | Chamarcrista rotundifolia | Senna hirsuta | Senna alata | Senna occidentalis | Senna siamea | Cassia fistula | Cassia sieberiana | Senna obtusifolia | Senna polyphylla | Senna surattensis |
| Stigma type | Lateral | Terminal | Involute | Lateral | Subtermin, punctiform |
Punctiform | Terminal/ subterminal |
Terminal | Subterminal | Lateral |
| Style apex | Glabrous | Short | Pruinose | Glabrous | Stout | Sturdy | Slender | Glabrous | Glabrous | Glabrous |
| Anther shape | Linear oblong | Curved/beaked | Curved, swollen and long |
Curved | Long and nearly straight | Long (longitudinal rim) |
Dorsifixed attachment |
Base knob like | Lateral ridged | Equal and straight with lateral longitudinal rim |
| Filament | Very short | Flat winged | Long | Long | Long | Long (towards the middle) |
Longer exceeding | Slender and glabrous the petal |
Curved | Thick and glabrous |
| Stamen | Unequal | Erect | Broadly elliptical | Erect | Glabrous | Curved | Unequal | Beaked and same size |
Curved | Thick glabrous |
| Pedicel | Filiform | Pubscent | Sparely pubescent | Pubscent | Sturdy | Elongated | Deflexed lower | Thinly pubescent | Long | Sparsely pubscent |
| Ovary | Ovary | Tomentose | Grooved | Appressed | Stipitate | Thinly appressed | Stipitate | Strigulate | Stipitate | Curved |
| Fruit colour | Blackish brown | Brown | Dark brown | Dark brown | Brown | Dark brown | Dark brown -black | Brown | Brown | Brown |
| Fruit shape | Linear oblong | Flat and angular (subterete) |
Winged and subterete |
Subterete and sickle shaped |
Flat | Terete | Straight and terete | Terete | Flat | Flat |
| Fruit surface | Glabrous | Ribbed hirsute | Chartaceous | Densely hirsute | Glabrous | Glabrous | Glabrous | Glabrous | Glabrous | Glabrous |
| Fruit dehiscence |
Elastically dehiscent valve coiling |
Dehiscent | Dehiscent | Dehiscent | Indehiscent | Indehiscent | Indehiscent | Dehiscent | Dehiscent | Dehiscent |
| Inflorescence type |
Racemose | Paniculate | Racemose | Paniculate | Paniculate | Racemose | Racemose | Racemose | Paniculate | Racemose |
| Inflorescence position |
Axillary | Axillary or rarely terminal |
Axillary | Axillary and terminal | Dense corymb | Axillary/terminal | Axillary | Axillary/ solitary | Axillary | Axillary |
| Seed colour | Pale brown | Dark olive | Brown | Brown | Light brown | Glossy brown | Brown | Glossy dark brown | Dark brown | Blackish smooth |
| Seed shape | Rhomboid to rectangular | Ovate | Obovate | Oval | Flat or oval | Flatten/transverse, ovate |
Ellipsoid | Septate/rhomboid | Oval | Flattened |
| Number of seeds per lobe |
Small seeds | 50-80 | 20-35 | 25-35 | 20-30 | 40-100 | Many | 20-50 | 10-20 | 20-35 |
| Table 4: | Quantitative macro-morphological characters of selected species in Cassiinae | |||
| Taxa | Leaflet length | Leaflet width | Rachis length | Petiolule length | Blade length | Length/width ratio | Blade/rachis ratio |
| Chamaecrista rotundifolia | 1.42±0.09 | 0.96±0.07 | 0.00±0.00 | 0.34±0.02 | 4.02±0.24 | 1:01 | N/A |
| Senna hirsuta | 5.01±0.38 | 2.28±0.14 | 12.04±0.79 | 0.11±0.01 | 10.98±0.69 | 3:02 | 6:07 |
| Senna alata | 13.62±2.66 | 6.61±0.80 | 25.53±2.52 | 0.18±0.03 | 30.37±3.75 | 7:04 | 16:13 |
| Senna occidentalis | 4.96±0.27 | 1.93±0.11 | 11.60±0.61 | 0.17±0.02 | 11.49±0.51 | 3:02 | 1:01 |
| Senna siamea | 4.67±0.89 | 1.79±0.13 | 15.89±5.20 | 0.23±0.03 | 11.13±1.76 | 3:01 | 3:04 |
| Cassia fistula | 10.13±0.31 | 5.13±0.13 | 24.97±6.23 | 0.53±0.01 | 24.68±0.78 | 2:01 | 1:01 |
| Cassia sieberiana | 4.89±0.59 | 2.39±0.28 | 15.77±2.51 | 0.41±0.04 | 11.93±1.18 | 3:02 | 3:04 |
| Senna obtusifolia | 2.33±0.63 | 1.36±0.40 | 4.52±0.75 | 0.18±0.03 | 5.49±1.77 | 2:01 | 1:01 |
| Senna polyphylla | 0.67±0.04 | 0.37±0.02 | 3.76±0.38 | 0.00 ±0.00 | 1.57±0.03 | 1:01 | 1:02 |
| Senna surattensis | 5.25±0.80 | 2.57±0.36 | 12.95±0.15 | 0.25±0.05 | 12.75±1.20 | 3:2 | 1:1 |
| Mean value±Standard Error of the Mean | |||||||
| Table 5: | Qualitative leaf micro-morphological characters of species in the Cassiinae | |||
| Cell shape | Stomata type | Anticlinal wall pattern | Cell wall striation | Trichome | Crystal | |||||||
| Taxa | AD | AB | AD | AB | AD | AB | AD | AB | AD | AB | AD | AB |
| Chamaecrista rotundifolia | Irregular | Irregular | Anomocytic | Anomocytic | Undulate | Undulate | - | - | + | + | - | - |
| Senna hirsuta | Irregular | Irregular | Anomocytic | Anisocytic | Straight and curved | Undulate | + | - | + | + | - | - |
| Senna alata | Irregular | Irregular | Anisocytic | Anomocytic | Undulate | Curved | - | + | + | - | - | - |
| Senna occidentalis | Polygonal | Polygonal | Anisocytic | Anisocytic | Straight and curved | Straight and curved | + | - | - | + | - | - |
| Senna siamea | Polygonal | Polygonal | Paracytic and anisocytic | Anisocytic | Straight | Straight | + | - | + | + | + | - |
| Cassia fistula | Polygonal | Polygonal | Absent | Absent | Straight | Straight | - | + | - | - | + | - |
| Cassia sieberiana | Polygonal | Polygonal | Absent | Anisocytic | Straight | Straight | - | - | - | - | - | - |
| Senna obtusifolia | Irregular | Irregular and polygonal | Anisocytic | Anisocytic | Curved | Curved | - | - | + | - | - | |
| Senna polyphylla | Irregular | Polygonal | Anisocytic and anomocytic |
Anisocytic and anomocytic |
Undulate | Straight | + | - | - | - | - | - |
| Senna surattensis | Polygonal | Irregular | anomocytic | Anisocytic | Straight | Straight and curved | - | - | + | + | + | - |
| AB: Abaxial, AD: Adaxial, +: Present and -: Absent | ||||||||||||
| Table 6: | Quantitative leaf micro characters of species in Cassiinae | |||
| Number of cells/×400 area of view |
Epidermal cell length (μm) | Epidermal cell width (μm) | Stomatal density per ×400 area of view |
Stomatal length (μm) | Stomatal width (μm) | S.I (%) | ||||||||
| OTUs | AD | AB | AD | AB | AD | AB | AD | AB | AD | AB | AD | AB | AD | AB |
| 1 | 10.75±3.99 | 125.25±9.08 | 28.93±1.90 | 31.25±1.72 | 23.13±0.82 | 25.10±0.53 | 53.25±5.18 | 45.25±4.43 | 1.70±0.05 | 4.94±0.66 | 1.33±0.05 | 1.28±0.03 | 81.31 | 66.17 |
| 2 | 140.25±11.76 | 222.75±8.44 | 29.75±1.56 | 30.00±1.86 | 25.75±1.29 | 22.75±1.02 | 39.00±3.34 | 126.00±7.91 | 2.48±0.04 | 2.40±0.08 | 1.68±0.11 | 1.88±0.16 | 55.91 | 90.14 |
| 3 | 191.75±6.71 | 291.25±9.62 | 23.25±1.97 | 16.00±1.19 | 22.25±1.84 | 14.00±0.41 | 79.00±3.60 | 172.00±12.44 | 2.20±0.06 | 2.18±0.15 | 1.40±0.18 | 1.20±0.06 | 75.49 | 91.88 |
| 4 | 164.50±8.10 | 218.00±9.85 | 29.75±1.60 | 31.25±1.41 | 26.50±1.07 | 30.25±1.26 | 67.75±4.75 | 119.75±5.87 | 2.08±0.13 | 2.13±0.10 | 1.35±0.06 | 1.20±0.05 | 72.86 | 88.76 |
| 5 | 222.25±14.12 | 298.00±50.65 | 28.25±1.35 | 26.25±2.30 | 21.25±1.76 | 19.25±2.08 | 19.50±6.13 | 45.50+8.69 | 2.03±0.10 | 2.18±0.09 | 1.3±0.06 | 1.33±0.07 | 20.42 | 40.94 |
| 6 | 340.0±23.31 | 314.75±9.14 | 23.00±1.33 | 24.25±1.58 | 13.25±0.38 | 15.25±1.20 | Absent | Absent | Absent | Absent | Absent | Absent | Absent | Absent |
| 7 | 338.25±17.45 | 381.00±17.87 | 13.00±0.73 | 18.75±2.02 | 12.00±0.33 | 11.25±0.42 | Absent | 12.00±1.66 | Absent | 2.45±0.09 | Absent | 1.10±0.06 | Absent | 7.68 |
| 8 | 156.25±14.21 | 191.50±15.18 | 30.75±1.45 | 4.89±1.55 | 19.75±1.15 | 24.00±1.00 | 55.25±4.87 | 42.00±7.91 | 1.23±0.08 | 1.80±0.10 | 0.35±0.04 | 0.17±0.06 | 66.51 | 43.19 |
| 9 | 256.25±0.31 | 248.50±0.20 | 25.25±0.06 | 24.75±0.16 | 10.00±0.09 | 23.75±0.14 | 66.75±0.22 | 79.75±0.39 | 2.35±0.15 | 2.13±0.19 | 1.1±0.13 | 1.23±0.14 | 52.6 | 61.27 |
| 10 | 192.25±13.65 | 280.75±12.55 | 28.75±1.08 | 30.25±2.05 | 25.25±0.73 | 27.50±1.55 | 9.25±2.47 | 118.50±4.65 | 2.73±0.08 | 2.50±0.14 | 2.13±0.09 | 1.40±0.06 | 12.02 | 74.85 |
| AB: Abaxial, AD: Adaxial, S.I (%): Stomatal index percentage and OTUs: Operational taxonomic units represents each species studied | ||||||||||||||
| Table 7: | Qualitative phytochemical characters of species in the Cassiinae | |||
| Taxa | SAP | ALK | FLA | TAN | COM | STD | TPD | CARD | PHL | QUN | GLY | PSD | ANTH |
| Chamaecrista rotundifolia | + | + | + | + | + | + | + | - | + | + | - | + | - |
| Senna hirsuta | + | + | + | + | + | + | + | - | + | + | - | + | - |
| Senna alata | + | + | + | + | + | + | + | - | + | + | + | + | - |
| Senna occidentalis | + | + | + | + | + | + | + | - | + | + | - | + | - |
| Senna siamea | + | + | + | + | + | + | + | + | + | + | - | + | - |
| Cassia fistula | + | + | + | + | + | + | + | - | + | + | - | + | - |
| Cassia sieberiana | + | + | + | + | + | + | + | - | + | + | - | + | - |
| Senna obtusifolia | + | + | + | + | + | + | + | - | + | + | - | + | - |
| Senna polyphylla | + | + | + | + | + | + | + | + | + | + | + | + | - |
| Senna surattensis | + | + | + | + | + | + | + | + | + | + | + | + | - |
| SAP: Saponins, ALK: Alkaloids, FLA: Flavonoids, TAN: Tannins, COM: Coumarins, STD: Steroids, TPD: Terpenoids, CARD: Cardiac glycosides, PHL: Phenols, QUN: Quinones, GLY: Glycosides, PSD: Phyto-steroids, ANTH: Anthraquinones, -: Absent and +: Present | |||||||||||||
| Table 8: | Quantitative phytochemical characteristics of species in Cassiinae | |||
| Taxa | Saponins (%) | Flavonoids (mg/g) | Alkaloids (%) | Tannins (mg/g) | Phenols (mg/g) |
| Chamaecrista rotundifolia | 2.57 | 26.78±0.38 | 17.05 | 3.20±0.07 | 6.76±0.04 |
| Senna hirsuta | 1.09 | 20.23±0.31 | 18.1 | 2.61±0.03 | 2.99±0.06 |
| Senna alata | 2.19 | 42.31±0.44 | 8.62 | 5.46±0.02 | 11.97±0.07 |
| Senna occidentalis | 1.55 | 34.05±0.38 | 10.95 | 4.32±0.03 | 4.99±0.04 |
| Senna siamea | 1.1 | 32.83±0.26 | 27.7 | 2.35±0.02 | 4.84±0.06 |
| Cassia fistula | 1.95 | 30.91±0.38 | 25.4 | 2.67±0.03 | 6.36±0.06 |
| Cassia sieberiana | 5.75 | 46.87±0.31 | 0.7 | 11.69±0.00 | 24.38±0.04 |
| Senna obtusifolia | 1.5 | 26.21±0.62 | 14.05 | 3.79±0.03 | 7.01±0.04 |
| Senna polyphylla | 3.25 | 32.19±0.38 | 10.45 | 7.13±0.04 | 14.84±0.03 |
| Senna surattensis | 4.9 | 34.47±0.31 | 21.4 | 10.40±0.00 | 19.26±0.04 |
Phytochemical analysis: Table 7 and 8 show the qualitative and quantitative phytochemical characters evaluated for species in the subtribe Cassinae. Results from qualitative analysis indicated the presence of phytochemical metabolites such as saponins, alkaloids, flavonoids, tannins, coumarins, steroids, terpenoids, phenols, quinones and phyto-steroids in all ten species studied. Other organic metabolites such as glycosides were present in Senna alata, Senna polyphylla and Senna surattensis. While cardiac glycosides were present in Senna Siamea, Senna polyphylla and Senna surattensis. The quantity of phytochemical metabolites examined ranges from highest to lowest in the following phytochemical compounds; saponins (5.75-1.09%)], alkaloids (25.40-0.70%), flavonoids (47.44-26.07 mg/g), tannins (11.69-2.31 mg/g) and phenols (4.74-24.44 mg/g).
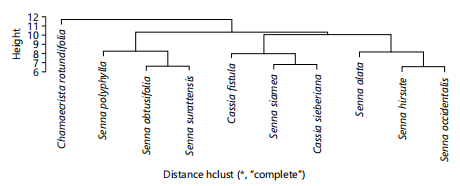
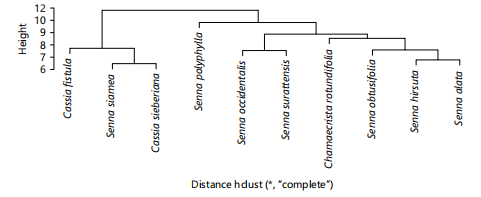
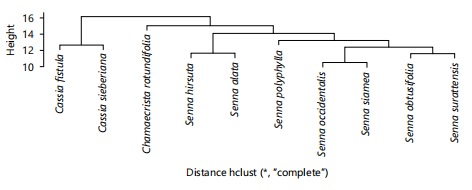
Cluster and principal component analysis: Figure 4 is dendrogram showing the phenetic relationship among studied species using morphological characters along four groups were produced. Group 1 contains Chamaecrista rotundifolia only, group 2 contains Senna polyphylla, S. obtusifolia and S. surattensis. Group 3 contains C. fistula, S. siamea and C. sieberiana, while group 4 contains S. hirsuta, S. alata and S. occidentalis. However, Fig. 5 shows phenetic relationship among species using average linkage analysis of anatomical characters. the dendrogram for anatomical characters produced for groups. Group one contains C. fistula, S. siamea and S. sieberiana. Group two contains S. Polyphylla only, group three contains S. occidentalis and S. surattensis, group four contains only C. rotundifolia while group five contains S. obtusifolia, S. hirsuta and S. alata. Figure 6 shows phenetic relationship of phytochemical characters. The dendrogram produced contains four groups. Group 1 (S. hirsuta, C. rotundifolia and S. obtusifolia), group 2 (S. siamea, S. occidentalis and C. fistula), group 3 (C. sieberiana) and group 4 (S. alata, S. polyphylla and S. surattensis). The dendrograms produced in Fig. 7 illustrates the clustering of 10×96 data matrix of all data evaluated in the three data sets (morphology, anatomy and phytochemistry). Three clusters were formed. Cluster 1 contains Cassia fistula and Cassia sieberiana. Cluster 2 contains Chamaecrista rotundifolia alone. Cluster 3 contains Senna hirsuta, Senna alata, Senna polyphylla, Senna occidentalis, Senna siamea, Senna obtusifolia and Senna surrattensis. The three clusters formed represent the three genera (Cassia, Senna and Chamaecrista).
|
|

|
|
|
| Table 9: | Squared Euclidean distance proximity matrix for 10×96 scored characters in ten species of Cassiinae | |||
| OTU | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| 2 | 13.30792 | ||||||||
| 3 | 15.01758 | 11.96474 | |||||||
| 4 | 12.56871 | 11.11334 | 12.02288 | ||||||
| 5 | 14.3244 | 12.16883 | 12.96011 | 10.61969 | |||||
| 6 | 16.85361 | 15.97632 | 15.38459 | 14.23571 | 11.62061 | ||||
| 7 | 17.19873 | 15.0766 | 14.93372 | 14.46792 | 11.63 | 13.11997 | |||
| 8 | 13.62632 | 13.12539 | 13.03021 | 12.25797 | 12.12502 | 15.27391 | 14.71405 | ||
| 9 | 15.05855 | 14.82971 | 14.38228 | 12.90279 | 12.97197 | 16.10664 | 14.65222 | 13.81268 | |
| 10 | 15.89423 | 14.22384 | 14.80636 | 12.85518 | 11.52693 | 15.06101 | 14.03293 | 11.93223 | 13.82951 |
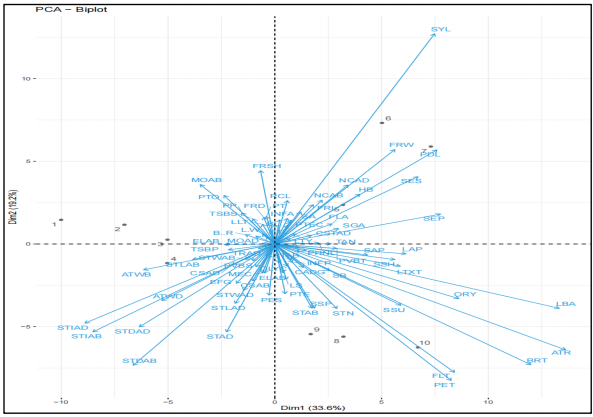
Figure 8 showing two-dimensional model of Principal Component Analysis (PCA) obtained from the ordination of principal component axis of I, II, III, IV for 96×10 morphological, anatomical and phytochemical data matrix. Numbers 1 to 10 correspond to OTUs as in Table 2. The OTUs that occurred in each of the axes of the chart include; axis I contains OTUs 5, 6 and 7 (Senna siamea, Cassia fistula and Cassia sieberiana), axis II contains OTUs 8, 9 and 10 (Senna obtusifolia, Senna polyphylla and Senna surattensis), axis III contains OTUs 4 (Senna occidentalis) and axis IV contains OTUs 1, 2 and 3 (Chamaecrista rotundifolia, Senna hirsuta and Senna alata). Although almost all OTUs in each axis were not in clusters, they still appeared on the same axis or group. The squared euclidean distance proximity matrix for 10×96 characters in ten species in the Cassiinae was shown in Table 9. The Eigen values, variance and cumulative variance percentage for ten character matrix was shown in Table 10 using the combined data sets from anatomical, morphological and phytochemical features. Eigen value was highest in OTU 5 (9.31) and variance percentage was highest in OTU 4 (9.50%), followed by OTU 5 (8.10%).
| Table 10: | Eigen values, variance and cumulative variance percentage for ten (10) OTU character matrix | |||
| OTU | Eigen value | Variance (%) | Cumulative variance (%) |
| 1 | 3.86E+01 | 3.36E+01 | 33.55286 |
| 2 | 2.21E+01 | 1.92E+01 | 52.76888 |
| 3 | 1.18E+01 | 1.03E+01 | 63.02281 |
| 4 | 1.09E+01 | 9.50E+00 | 72.52192 |
| 5 | 9.32E+00 | 8.10E+00 | 80.62228 |
| 6 | 7.36E+00 | 6.40E+00 | 87.02477 |
| 7 | 5.82E+00 | 5.06E+00 | 92.08324 |
| 8 | 4.97E+00 | 4.32E+00 | 96.40352 |
| 9 | 4.14E+00 | 3.60E+00 | 100 |
| 10 | 2.53E-30 | 2.20E-30 | 100 |
| Table 11: | Key to the species in the subtribe Cassiinae in Nigeria | |||
| 1a | Herb shrub or tree with leaves pari pinnately compound, inflorescence raceme, venation pinnate, margin, entire, leaflet arrangement alternate…………………………………………………….……………..……………………1 |
| 2 | Trees woody, with more than 10 m in height, bracteole present, leaf base rounded to broadly cuneate, leaf texture coriaceous, fruit indehiscent, leaf apex acute……………………………………………………..…1. Cassia fistula |
| 2 | Leaf base rounded to cuneate, leaf texture coriaceous, fruit indehiscent leaf apex subacute ……………………………………… …………...………………....,,,…..…............................…2. Cassia sieberiana |
| 3 | Tree or shrub with less than 10 m in height with woody or herbaceous stem……………………...………………...3 |
| 4 | Bract shape obovate, bracteole absent, pedicel sparsely pubescent, cardiac glycosides present…………………………………… …………………………………………..………………3. Senna siamea |
| 4 | Bract shape obovate, bracteole absent, pedicel sparsely pubescent cardiac glycosides absent ……….……………………………….… ………………………………………………………...……4. Senna alata |
| 5 | Shrub with over 5 m in height………………………………………………………….…………………………….5 |
| 6 | Glycosides present, anticlinal wall pattern straight, inflorescence position fascicle ……………………………………………………...……… …..………….................................…5. Senna polyphylla |
| 6 | Glycosides present, anticlinal cell wall pattern straight and curved inflorescence position solitary……………………………………………………………………………….......….........6. Senna surattensis |
| 7 | Herb equal to 2 m in height herbaceous……………………………………..….……………………………………7 |
| 8 | Stipule shape linear, stigma terminal, seed shape rhomboidal, epidermal cell shape irregular leaf surface above villous ……………………………………………………………………………..…………………7. Senna hirsuta |
| 8 | Stipule shape linear, stigma terminal, seed shape rhomboidal, epidermal cell shape irregular, leaf surface above glabrous………………………………….………………………..………………………...….…8. Senna obtusifolia |
| 1b | Shrub or herb with leaves pinnately compound, inflorescence raceme, venation pinnate, margin entire, leaflet arrangement alternate…………………………...…………………………………………...……………………….9. |
| 9 | Shrub or herb equal or less than 2 m height ……………………………………………………...…...………….….9 |
| 10 | Petal shape obovate, dehiscent, fruits, pedicel pubescent, …………..………………...…….…9. Senna occidentalis |
| 10 | Petal shape obovate, elastically dehiscent valve coiling fruit, pedicel filiform, ……………………………………...…………………………… …………….……10. Chamaecrista rotundifolia |
The least value was obtained in OTU 3 as 1.03%. Cumulative variance was highest in OTUs 9 and 10, weighing a 100% each. Table 11 shows the key to the identification of species constructed based on characters of OTU evaluated in this work. It is important to create a very simple taxonomic key for identification and better understanding of shared or apomorphic characters.
DISCUSSION
The morphological, anatomical and phytochemical characters used for this study revealed the extent of variation and proved to be taxonomically relevant in the delimitation of selected taxa. The combination of characters from floral, vegetal, anatomy and phytochemical metabolites is employed to better understand the phenetic relationship among members of the subtribe Cassiinae. In Table 4-6 the dendrograms produced confirmed that taxonomic classification approaches do not rely on just morphological data but need to be used in combination with a wide variety of characters from different scientific fields to delimit and classify a particular taxon. Morphological data has been used successfully in combination with pollen characters to delimit the subtribe Cassinae19. The most informative morphological data sets identified in this work produced a great range of similarities. The PCA identified distinctiveness characters such as stomata index, stomata density, filament length, petal shape, bract shape, nature of ovary, leaf base and anther contributing about 33.6% to the delimitation, while style apex, fruit width, pedicel, sepal surface and sepal shape contributed about 19.2%. The results in this work correspond with research on important morphological characters in several species of Cassiinae in South-Western Nigeria20. It is of interest to note that each data set analyzed separately which includes morphology, anatomy and phytochemical characters was not able to delimit the subtribe in isolation as shown in Fig. 4-6. In this study cluster analysis and principal component analysis are two component analytical tools used to trace relationships among members of the Cassiinae clade. The revelation of marked differences in the dendrogram produced for morphological, anatomical and phytochemical characters assessed is a strategic indication that one data character alone is insufficient to make a taxonomic conclusion that a particular group shares the same ancestral traits. The combination of multiple data characters was essential in the delimitation and classification of the groups. A similar approach was obtained in the numerical taxonomy of species of the subtribe Cassiinae reported by Kolawole et al.21. The difference in the clusters produced for phytochemical characters is a result of the different chemical affinity among studied taxa22. Results obtained from the combined data for morphology, anatomy and phytochemical characters produced a dendrogram that agreed with the taxonomic treatment of the subtribe by Irwin and Barneby8 separating the species into distinct genera. This implies Cassia species, Senna species and Chamaecrista species are distinct genera and should be treated as such consequently supporting the polyphyletic nature of the subtribe. Principal Component Analysis (PCA) shows the following morphological characters contributed more to the closeness or affinity in the relationship among taxa petal shape, fruit length, bract, anther, leaf base, style and pedicel. The results also supported the report by the Legume Phylogeny Working Group on the Legume phylogeny and classification in the 21st century23. Hence qualitative floral morphological characters contributed more to delimitation and relationship among groups in this study. In the case of anatomical characters, the PCA analysis reveals that the anticlinal walls on both adaxial and abaxial surfaces, stomata density on adaxial and abaxial surfaces, stomata indices on adaxial and abaxial surfaces and petiolule vascular bundle shapes were a contributor to the variation among the taxa. The PCA analysis for phytochemical characters revealed that saponins were a major contributor to similarity in the group. Thus, phytochemical metabolic variations are significant characters in the classification of plants.
CONCLUSION
This research therefore has confirmed the autonomy and classifications of members of the subtribe Cassiinae into three generic categories using a combined data set which serves as a reliable database for natural classification of members of the group and suggests further findings by combining gross morphological and molecular data sets. This study has affirmed the importance of using multiple data sets to delimit members of a taxonomic clade. It is clear from the dendrogram produced for combined 10×96 data matrix for morphology, anatomy and phytochemical features that the systematic combination and analysis of data could serve as potentially significant criteria for identification but not a reliable delimitation tool when used in isolation for classification. The combination of the three data sets produced corresponding results similar to previous works on the subtribes and formed a reliable basis for an identification key.
SIGNIFICANCE STATEMENT
This study aimed to determine the taxonomic relationships among selected species in the subtribe cassiinae using the numerical taxonomic principles. The research objectives are focused on the evaluation of morphological and phytochemical character variations and determination of phenetic relationships among selected species in Cassiinae taxonomic key produced was relevant in the identification of the studied group. The findings in this work affirmed the importance of using multiple data sets to delimit members of a taxonomic clade. The combination of data sets from morphological, anatomical and phytochemical characters served as a reliable database for identification and natural classification. Further findings aimed to compliment and improve the traditional method of taxonomic delimitation used in this work is recommended.
REFERENCES
- Lewis, G.P., B.D. Schrire, B.A. Mackinder, L. Rico and R. Clark, 2013. A 2013 linear sequence of legume genera set in a phylogenetic context-A tool for collections management and taxon sampling. South Afr. J. Bot., 89: 76-84.
- Yahara, T., F. Javadi, Y. Onoda, L.P. de Queiroz and D.P. Faith et al., 2013. Global legume diversity assessment: Concepts, key indicators, and strategies. TAXON, 62: 249-266.
- Eseyin, O.A., U. Benedict, P.S. Thomas, I. Etim and E. Essien et al., 2018. Isolation and characterization of antioxidant constituents of the fruit of Telfairia occidentalis Hook F (Cucurbitaceae). Trop. J. Pharm. Res., 17: 1953-1960.
- Koenen, E.J.M., 2022. Osodendron gen. nov. (Leguminosae, Caesalpinioideae), a new genus of mimosoid legumes of tropical Africa. PhytoKeys, 205: 453-470.
- Davis, P.H. and V.H. Heywood, 1973. Principles of Angiosperm Taxonomy. Krieger, Florida, United State, Pages: 558.
- Wojciechowski, M.F., M. Lavin and M.J. Sanderson, 2004. A phylogeny of legumes (Leguminosae) based on analysis of the plastid matK gene resolves many well-supported subclades within the family. Am. J. Bot., 91: 1846-1862.
- Lock, J.M., 1988. Cassia Sens. Lat. (Leguminosae-Caesalpinioideae) in Africa. Kew Bull., 43: 333-342.
- Irwin, H.S. and R.C. Barneby, 1982. The American Cassiinae: A Synoptical Revision of Leguminosae Tribe Cassieae Subtribe Casiinae in the New World. New York Botanical Garden, New York, ISBN: 9780893272418, Pages: 918.
- Ayodele, A.E and J.D. Olowokudejo, 2006. The family Polygonaceae in West Africa: Taxonomic significance of leaf epidermal characters. South Afr. J. Bot., 72: 442-459.
- Chukwuma, E.C., A.E. Ayodele, M.O. Soladoye and D.M. Adewumi, 2016. Numerical taxonomy of Berlinia species (Caesalpinioideae: Leguminosae) and their distribution in Nigeria. J. Threatened Taxa, 8: 8443-8451.
- Banji, A., B. Goodluck, O. Oluchi and F. Stephen, 2018. Antimicrobial and antioxidant activities of crude methanol extract and fractions of Andrographis paniculata leaf (Family: Acanthaceae) (Burm. f.) Wall. Ex Nees. Jordan J. Biol. Sci., 11: 23-30.
- Obadoni, B.O. and P.O. Ochuko, 2002. Phytochemical studies and comparative efficacy of the crude extracts of some haemostatic plants in Edo and Delta States of Nigeria. Global J. Pure Appl. Sci., 8: 203-208.
- Mythili, K., C.U.M. Reddy, D. Chamundeeswari and P.K. Manna, 2014. Determination of total phenol, alkaloid, flavonoid and tannin in different extracts of Calanthe triplicate. Res. Rev.: J. Pharmacogn. Phytochem., 2: 40-44.
- Sheba, L.A. and A. Venkatraman, 2021. Physicochemical characterization, phytochemical and HPTLC fingerprinting studies on fruit of Couroupita Guianensis. Chiang Mai Univ. J. Nat. Sci., 20.
- Hikino, H., Y. Kiso, H. Wagner and M. Fiebig, 1984. Antihepatotoxic actions of flavonolignans from Silybum marianum fruits. Planta Med., 50: 248-250.
- Ejikeme, C.M., C.S. Ezeonu and A.N. Eboatu, 2014. Determination of physical and phytochemical constituents of some tropical timbers indigenous to Niger Delta area of Nigeria. Eur. Sci. J., 10: 247-270.
- Jayapriya, G. and F.G. Shoba, 2014. Screening for phytochemical activity of Urechites lutea plant. Asian J. Plant Sci. Res., 4: 20-24.
- Sokal, R.R. and P.H.A. Sneath, 1963. Principles of Numerical Taxonomy. W.H. Freeman, New York, ISBN: 9780716706212, Pages: 359.
- Saheed, S.A. and H.C. Illoh, 2010. A taxonomic study of some species in Cassiinae (Leguminosae) using leaf epidermal characters. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 38: 21-27.
- Saheed, S.A. and H.C. Illoh, 2011. Important morphological characters in several species of Cassiinae (Leguminosae) in South-Western Nigeria. Notulae Sci. Biol., 3: 47-56.
- Kolawole, O.S., A.A. Abdulrahaman, E.C. Chukwuma and M.A. Jimoh, 2021. A numerical approach to the taxonomy of some species of the subtribe Cassiinae in Nigeria. Webbia: J. Plant Taxon. Geogr., 76: 97-108.
- Bruneau, A., F. Forest, P.S. Herendeen, B.B. Klitgaard and G.P. Lewis, 2001. Phylogenetic relationships in the Caesalpinioideae (Leguminosae) as inferred from chloroplast trnL intron sequences. Syst. Bot., 26: 487-514.
- Legume Phylogeny Working Group, A. Bruneau, J.J. Doyle, P. Herendeen and C. Hughes et al., 2013. Legume phylogeny and classification in the 21st century: Progress, prospects and lessons for other species-rich clades. TAXON, 62: 217-248.
How to Cite this paper?
APA-7 Style
Onyinyechi,
O., Ayodele,
A. (2024). Morphologicaand Phytochemical Characterization of Cassiinae in Nigeria. Trends in Agricultural Sciences, 3(3), 274-295. https://doi.org/10.17311/tas.2024.274.295
ACS Style
Onyinyechi,
O.; Ayodele,
A. Morphologicaand Phytochemical Characterization of Cassiinae in Nigeria. Trends Agric. Sci 2024, 3, 274-295. https://doi.org/10.17311/tas.2024.274.295
AMA Style
Onyinyechi
O, Ayodele
A. Morphologicaand Phytochemical Characterization of Cassiinae in Nigeria. Trends in Agricultural Sciences. 2024; 3(3): 274-295. https://doi.org/10.17311/tas.2024.274.295
Chicago/Turabian Style
Onyinyechi, Okpara, and Abiodun Ayodele.
2024. "Morphologicaand Phytochemical Characterization of Cassiinae in Nigeria" Trends in Agricultural Sciences 3, no. 3: 274-295. https://doi.org/10.17311/tas.2024.274.295

This work is licensed under a Creative Commons Attribution 4.0 International License.