Histopathology of Root-Knot Nematode (Meloidogyne incognita) Infection on Two African Indigenous Vegetables
-
Leonard Uzoma Amulu


Department of Crop Science, Faculty of Agriculture, University of Agriculture and Environmental Sciences, Umuagwo, Nigeria
Durdoluwa Joseph OyedeleDepartment of Soil Science and Land Resources Management, Faculty of Agriculture, Obafemi Awolowo University, Ile-Ife, Nigeria
Ojo Kolawole AdekunleDepartment of Crop Production and Protection, Faculty of Agriculture, Obafemi Awolowo University, Ile-Ife, Nigeria
| Received 09 Mar, 2024 |
Accepted 29 Jun, 2024 |
Published 30 Jun, 2024 |
Background and Objective: The root-knot nematodes are cosmopolitan pests attacking almost all cultivated crops including African vegetables. Two screen house studies were conducted in 2016 to investigate the histopathology of Meloidogyne incognita on Solanum macrocarponandAmaranthus cruentus. Materials and Methods: The roots of 40 seedlings of Solanum macrocarponwere inoculated with 1,000 eggs of Meloidogyne incognita; the remaining 40 seedlings served as controls. Ten inoculated and 10 uninoculated seedlings were uprooted at seven days intervals. The study was repeated once. The roots were processed and sectioned. The sections were stained and examined under a compound microscope to determine giant cell formation. The establishment of Amaranthus cruentustrial followed a similar pattern to that of Solanum macrocarpon. Results: The Solanum macrocarpon and Amaranthus cruentus are susceptible to Meloidogyne incognita.This was evidenced by the formation of giant cells in the root tissues of the two vegetables. Conclusion: Solanum macrocarpon and Amaranthus cruentus are susceptible to Meloidogyne incognita.
| Copyright © 2024 Amulu et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
African indigenous vegetables are highly nutrient-dense and as such they supply food in the form of vitamins, mineral elements and antioxidants1. They are major sources of nutritious food for the poor rural families in Nigeria, who lack the resources to purchase high-value food such as egg, meat and milk. They contain high levels of vitamins and minerals which are known to be deficient among poor populations in Sub-Saharan African Countries including Nigeria2. They have great potential to improve food security and income amongst farming communities3. The relevance of these vegetables in balancing dietary quality is apparent in the traditional combination of starchy staples with a green vegetable sauce4.
Besides their nutritional qualities, they are rich in medicinal properties and are traditionally used in the treatment and management of various ailments such as constipation, cardiovascular diseases and diabetes1-5. A combination of these and many more has made these vegetables gain an important recognition as a solution to food insecurity, malnutrition and poverty6. Solanum macrocarpon is a commonly grown Solanum species in the humid tropics of Africa. They are important fruity or leafy vegetables in this region. They are rich in calcium, potassium, iron and magnesium. They are used for the treatment of many ailments such as measles, abdominal worms and stomach-aches in children and to ease delivery in pregnant women1.
Amaranthus cruentus is a nutritious vegetable with edible leaves and seeds. The leaves and seeds have been identified as good sources of vitamin A for children7. The protein component of the seeds has been shown to contain higher concentrations of lysine and sulfur compared to that contained in cereal grains and grain legumes, respectively.
The production of the vegetables is on increase as a result of the increased knowledge of their importance in fighting malnutrition, hunger and poverty, however, actual yields are very much lower than realized yields in many parts of Africa and this is partly due to poor soils and high levels of pests and diseases8,9.
Root-knot nematode, Meloidogyne incognita, is a cosmopolitan pest10. It attacks almost all cultivated crops causing a significant reduction in yields and quality of susceptible crops11,12. The nematode is recognized as the most devastating pest affecting vegetables in the tropical and subtropical regions of the world10.
Their feeding habit induces the formation of galls on the roots of infected plants, thereby, reducing the efficiency by which the plant root system performs its normal functions12. The mechanisms by which this nematode causes disease in plants have been reported on a number of exotic vegetables, however, there are little or no reports of this on many African indigenous vegetables including A. cruentus and S. macrocarpon, hence, this study investigated the histopathology of root-knot nematode (Meloidogyne incognita) infection on S. macrocarpon and A. cruentus.
MATERIALS AND METHODS
Study area: The experiment was conducted at the screen house of the Faculty of Agriculture, Obafemi Awolowo University, Ile-Ife, Nigeria in 2018.
Planting operations and inoculation: The 81 L plastic pots were filled with steam sterilized topsoil and these were arranged in 10×8 rows on screen house benches. Two weeks old seedlings of S. macrocarpon were transplanted into the pots at the rate of two seedlings per pot. The 2 weeks after transplanting, 1000 eggs of M. incognita in water suspension were inoculated close to the roots of each S. macrocarpon seedlings in each of 40 pots with the aid of an Eppendorf pipette. Soils from the pot were used to cover the roots immediately after inoculation was done, while distilled water was poured around the root systems of the remaining 40 seedlings (controls). The seedlings wetted as when due. The temperature of the the screen house and soil temperature of the potted plants were monitored; at 12:00 noon and 6:00 pm every day. The establishment of A. cruentus trial followed a similar trend to those of S. macrocarpon and both experiments were repeated once without any modifications.
Examination of giant cells and nematode development in roots of the vegetables: The 7 days after inoculation (DAI), 10 inoculated and 10 uninoculated (control plants) S. macrocarpon seedlings were randomly and carefully uprooted and subsequently at 14, 21 and 28 DAI. The roots of the seedlings were washed in gentle running water to remove adhering soils, the roots were dried between facial tissues and cut into smaller pieces of about 2 cm. The root pieces were put in fixative bottles and fixed in 50% ethanol. The fixed root segments were dehydrated in graded ethanol series of 60, 70, 90 and 100% at 2 hrs intervals. Then they were dehydrated in a series of ethanol and xylene solutions and embedded in paraffin wax. Serial longitudinal sections 8-10 μm thick were cut with a rotary microtome into a hot water bath, where floating sections were picked up with glass slides pre-coated with haupt’s adhesive. Each slide was de-waxed in xylene, stained in Safranin O and counter-stained in fast green13. All sections on the glass slides were covered with cover slips and sealed with Canadian balsam, prior to examination with a compound microscope (Motic microscopes; model: 61400131). Observations were made on the integrity of cellular structures, nematode development and formation of giant cells and photomicrographs were taken after observation.
RESULTS
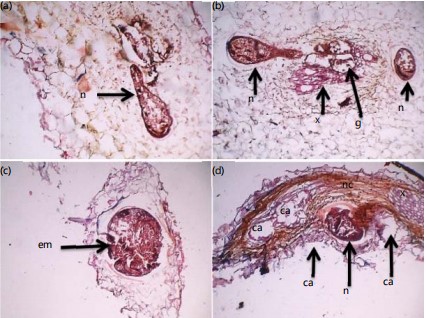
The mean ambient temperatures of the screen house at 12 noon and 6.0 pm were 32 and 30°C, respectively. The mean temperatures of soil at 12 noon and 6.0 pm were 32 and 29°C, respectively. Microscopic examinations of thin sections of roots of inoculated Solanum macrocarpon revealed the presence of fourth stage juveniles of M. incognita, distorted vascular tissues, giant cells around the head of the nematode and compressed cells (Fig. 1a and b). Egg mass was found in the root section of the vegetable at 28 DAI as shown in (Fig. 1c). There was adult female of M. incognita, distorted xylem tissues, necrotic lesions, small and large cavities and swollen epidermal cells in root sections of the vegetable at 28 DAI (Fig. 1d).
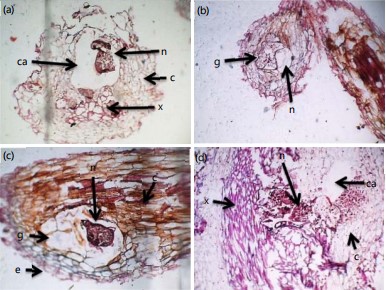
Examinations of thin sections of inoculated A. cruentus roots show the presence of fourth-stage juveniles, distorted vascular tissues, cortical cells and cavities within the cortical cells (Fig. 2a). There were fourth stage juvenile and giant cells around the head of the nematode in root tissues of A. cruentus at 21 DAI (Fig. 2b). Microscopic examinations of the root sections of A. cruentus at 28 DAI, revealed the presence of an adult female of M. incognita, giant cells aroundF the head of the nematode, compressed cortical cells and distorted xylem tissues (Fig. 2c). There were adult females of the nematode, large cavities within cortical cells and distorted xylem tissues in root section of the vegetable at 28 DAI (Fig. 2d).
|
|
|
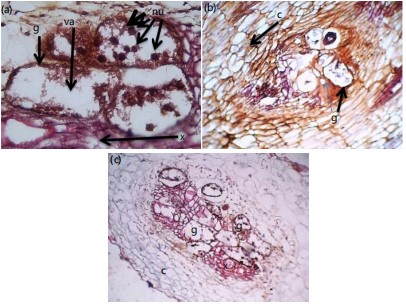
Examinations of root tissues of S. macrocarpon revealed the presence of distorted xylem tissue, giant cells containing vacuoles and numerous nuclei at 28 DAI (Fig. 3a). Figure 3b revealed the presence of compressed cortical cells and giant cells containing nuclei and vacuole in root section of A. cruentus. There were giant cells around a distorted vascular system in root section of S. macrocarpon at 21K.
The DAI I is section of inoculated S. macrocarpon (arrows point at giant cells (g), vacuole (va) and nucleus (nu)) at 21 DAI. The J is section of inoculated Amaranthus cruentus (arrows point at cortical cells (c) and giant cells (g) at 21 DAI. The K is section of inoculated S. macrocarpon (giant cells (g), distorted xylem (x) and cortical cells (c) (Fig. 3c).
DISCUSSION
The results of this study showed that M. incognita penetrated roots of S. macrocarpon and A. cruentus induced the formation of giant cells and consequently disorganized the cellular structures of root tissues. The reactions of these vegetables following nematode invasion were similar to those reported in roots of other susceptible crops14,15. Microscopic observations of thin sections of the two vegetables revealed the presence of pre-adult stage, adult stage and egg mass of the nematode, indicating that the nematode penetrated the host cell and migrated through the cortical region to the target cells where they established feeding sites. Once feeding site is established sedentary lifestyle commences as the nematode begins to obtain nutrients and water and reproduce.
The development and reproduction depend on the establishment and maintenance of feeding sites within the root tissues; hence nematodes depend on feeding sites for food and water to maintain and complete their entire life cycle. As the nematode feeds, it induces the formation of giant cells, although it is not known how these giant cells are produced, glandular secretions produced by the nematode and injected into plant cells through the stylet are suspected to interact directly or indirectly with the plant nuclear genome, thereby altering the gene expression that results in induction of giant cells14,16,17. The location of the giant cells around the head of the nematodes in this study and other similar studies support this fact18,19.
Many histological studies revealed the presence of giant cells close to the vascular tissues18,20,21. Microscopic observations of thin sections of the vegetables in this study also revealed similar observations as all giant cells observed were located close to the vascular tissues. This is an indication that these cells are metabolic sinks that direct photosynthates and water from leaves and root systems respectively to the feeding sites of the nematode20-22.
Giant cells are known to divide and expand in size with time without restrictions through the process known as hyperplasia and hypertrophy23. Microscopic observations of thin sections of several susceptible crop cultivars have revealed an increase in sizes of giant cells on a number of root-knot nematode-infected root tissues18. It is possible that neighboring cells and cortical cells are crushed following expansion and enlargement of giant cells.
In the present study, there were collapsed and disorganized cellular structures, particularly at 28 DAI. Apparently, as these cells expand the neighboring cells are compressed, crushed and pushed outward, resulting in the collapse and disorganization of the cellular structures observed in this study. Compression of neighboring cells, necrotic lesions, large cavities, collapse and disorganizations of cellular structures including xylem tissues following an increase in the size of giant cells have been reported by earlier workers19-24. Cabasan et al.15 reported collapsed structural features, thickened cortex and necrotic lesions by root-knot nematode M. graminicola in thin sections of root tissues of a susceptible rice cultivar. Similarly, Akhtar and Hisamuddin19 reported that microscopic examinations of thin sections of Vigna mungo infected by M. incognita revealed disruptions in the arrangement of vascular and cortical tissues within 25 and 30 DAI following expansions of giant cells.
The aftermath effects of disfigured cellular structures of plants include disruptions of the free flow of photosynthates and water from phloem and xylem tissues respectively to parts of the plants where they are needed. Distorted and galled root systems and yield reductions are usually the symptoms of plants heavily infected with the nematode. Many authors have reported a reduction in yield and quality of a number of vegetables following root-knot nematode infection in both field and screenhouse trials11-12. It is therefore important to control the activities of this parasite on fields cultivated to S. macrocarpon and A. cruentus in order to sustain yields.
CONCLUSION
The histopathology of Solanum macrocarpon and Amaranthus cruentus revealed that the vegetables are susceptible to Meloidogyne incognita. This was evidenced by the presence of different stages of the nematode life cycle (fourth stage juveniles, adult females and egg mass), in the root tissues of the plants at 21 and 28 DAI, which indicates development and positive relationship between the parasite and the nematode. The results of this finding suggest that Solanum macrocarpon and Amaranthus cruentus are susceptible to Meloidogyne incognita.
SIGNIFICANCE STATEMENT
Meloidogyne incognita is problematic in vegetable fields and has been reported to cause reductions in yields of vegetables. Their feeding habit induces the formation of galls on the roots of plants and slows down the movement of water and nutrients through the roots to other parts of the plant. Histopathology studies reveal the mechanism by which this nematode causes disease in crop plants. However, this has been studied on a number of exotic vegetable crops but little or no information is known on the histopathology of the nematode on African indigenous vegetable crops. Hence the study investigated the histopathology of Meloidogyne incognita on Solanum macrocarpon and Amaranthus cruentus. The result showed that Meloidogyne incognita penetrated the roots of the two vegetables and induced the formation of giant cells on the vegetables.
ACKNOWLEDGEMENTS
This research was supported by International Development Research Centre and the Department of Foreign Affairs, Trade and Development/Canadian International Food Security Research Fund through Project 107983 on synergizing indigenous vegetables and fertilizer micro-dosing innovations among West African farmers.
REFERENCES
- Schippers, R.R., 2000. African Indigenous Vegetables: An Overview of the Cultivated Species. Natural Resources Institute, Chatham, United Kingdom, ISBN: 0859545156, Pages: 214.
- Ishiekwene, I.C., T.E. Dada, J. Odoko and E.U. Nwose, 2019. Promoting African indigenous vegetables and its medical nutrition properties: A mini-narrative review based on Ukwani communities of Delta State Nigeria. Integr. Food Nutr. Metab., 6.
- Yiridoe, E.K. and V.M. Anchirinah, 2005. Garden production systems and food security in Ghana: Characteristics of traditional knowledge and management systems. Renewable Agric. Food Syst., 20: 168-180.
- Gido, E.O., O.I. Ayuya, G. Owuor and W. Bokelmann, 2017. Consumption intensity of leafy African indigenous vegetables: Towards enhancing nutritional security in rural and urban dwellers in Kenya. Agric. Food Econ., 5.
- Hilou, A., O.G. Nacoulma and T.R. Guiguemde, 2006. In vivo antimalarial activities of extracts from Amaranthus spinosus L. and Boerhaavia erecta L. in mice. J. Ethnopharmacol., 103: 236-240.
- Ambrose-Oji, B., 2009. Urban Food Systems and African Indigenous Vegetables: Defining the Spaces and Places for African Indigenous Vegetables in Urban and Peri-Urban Agriculture. In: African Indigenous Vegetables in Urban Agriculture, Shackleton, C.M., M.W. Pasquini and A.W. Drescher (Eds.), Routledge, England, United Kingdom, ISBN: 9781849770019, pp: 1-33.
- Akhtar, S., C. Karak, P. Biswas, A. Chattopadhyay and P. Hazra, 2012. Indigenous leafy vegetables: A potential source of β-carotene and ascorbic acid. Int. J. Veg. Sci., 18: 370-375.
- Adekunle, O.K., B.J. Amujoyegbe, M.K. Idowu and D.J. Oyedele, 2015. Incidence and management of plant parasitic nematodes under continuous vegetable production in a rainforest agroecology in Nigeria. J. Hortic. Sci. Biotechnol., 90: 20-24.
- Ogundele, R.A., D.J. Oyedele and O.K. Adekunle, 2016. Management of Meloidogyne incognita and other phytonematodes infecting Amaranthus cruentus and Telfairia occidentalis with African marigold (Tagetes erecta) and Siam weed (Chromolaena odorata). Australas. Plant Pathol., 45: 537-545.
- Sikora, R.A. and E. Fernández, 2005. Nematode Parasites of Vegetables. In: Plant Parasitic Nematodes in Subtropical and Tropical Agriculture, Luc, M., R.A. Sikora and J. Bridge (Eds.), CABI Publishing, Wallingford, England, ISBN: 978-1-84593-144-5, pp: 319-392.
- Niyaz, T., T. Azam and Hisamuddin, 2011. Histological responses and damage potential in roots of Eclipta alba L caused by Meloidogyne incognita. Libyan Agric. Res. Center J. Int., 2: 118-122.
- Amulu, L.U. and O.K. Adekunle, 2015. Comparative effects of poultry manure, cow dung, and carbofuran on yield of meloidogyne incognita-infested okra. J. Agric. Sci. Technol., 17: 495-504.
- Ravichandra, N.G., 2010. Methods and Techniques in Plant Nematology. PHI Learning Pvt. Ltd., Delhi, India, ISBN: 9788120340961, Pages: 616.
- Proite, K., R. Carneiro, R. Falcão, A. Gomes, S. Leal-Bertioli, P. Guimarães and D. Bertioli, 2008. Post‐infection development and histopathology of Meloidogyne arenaria race 1 on Arachis spp. Plant Pathol., 57: 974-980.
- Cabasan, M.T.N., A. Kumar, S. Bellafiore and D. de Waele, 2014. Histopathology of the rice root-knot nematode, Meloidogyne graminicola, on Oryza sativa and O. glaberrima. Nematology, 16: 73-81.
- Vanholme, B., J. de Meutter, T. Tytgat, M. van Montagu, A. Coomans and G. Gheysen, 2004. Secretions of plant-parasitic nematodes: A molecular update. Gene, 332: 13-27.
- Grunewald, W., M. Karimi, K. Wieczorek, E. van de Cappelle and E. Wischnitzki et al., 2008. A role for AtWRKY23 in feeding site establishment of plant-parasitic nematodes. Plant Physiol., 148: 358-368.
- Seo, Y., J. Park, Y.S. Kim, Y. Park and Y.H. Kim, 2014. Screening and histopathological characterization of korean carrot lines for resistance to the root-knot nematode Meloidogyne incognita. Plant Pathol. J., 30: 75-81.
- Akhtar, A. and Hisamuddin, 2015. Studies on the histopathological responses of Vigna mungo towards Meloidogyne incognita at different time intervals. J. Plant Pathol. Microbiol., 6.
- Caillaud, M.C., G. Dubreuil, M. Quentin, L. Perfus-Barbeoch and P. Lecomte et al., 2008. Root-knot nematodes manipulate plant cell functions during a compatible interaction. J. Plant Physiol., 165: 104-113.
- Abad, P., P. Castagnone-Sereno, M.N. Rosso, J. de A. Engler and B. Favery, 2009. Invasion, Feeding and Development. In: Root-Knot Nematodes, Perry, R.N., M. Moens and J.L. Starr (Eds.), CABI International, Wallingford, England, ISBN: 978-1-84593-492-7, pp: 163-181.
- Bartlem, D.G., M.G.K. Jones and U.Z. Hammes, 2014. Vascularization and nutrient delivery at root-knot nematode feeding sites in host roots. J. Exp. Bot., 65: 1789-1798.
- Ravichandra, N.G., 2013. Fundamentals of Plant Pathology. PHI Learning Private Limited, New Delhi, India, ISBN: 9788120347038, Pages: 652.
- Cabrera, V.A., M.E. Doucet and P. Lax, 2023. Histopathology of the root-knot nematode, Meloidogyne incognita, on ornamental plants (Crassulaceae). J. Plant Dis. Prot., 130: 891-897.
How to Cite this paper?
APA-7 Style
Amulu,
L.U., Oyedele,
D.J., Adekunle,
O.K. (2024). Histopathology of Root-Knot Nematode (Meloidogyne incognita) Infection on Two African Indigenous Vegetables. Trends in Agricultural Sciences, 3(2), 100-106. https://doi.org/10.17311/tas.2024.100.106
ACS Style
Amulu,
L.U.; Oyedele,
D.J.; Adekunle,
O.K. Histopathology of Root-Knot Nematode (Meloidogyne incognita) Infection on Two African Indigenous Vegetables. Trends Agric. Sci 2024, 3, 100-106. https://doi.org/10.17311/tas.2024.100.106
AMA Style
Amulu
LU, Oyedele
DJ, Adekunle
OK. Histopathology of Root-Knot Nematode (Meloidogyne incognita) Infection on Two African Indigenous Vegetables. Trends in Agricultural Sciences. 2024; 3(2): 100-106. https://doi.org/10.17311/tas.2024.100.106
Chicago/Turabian Style
Amulu, Leonard, Uzoma, Durdoluwa Joseph Oyedele, and Ojo Kolawole Adekunle.
2024. "Histopathology of Root-Knot Nematode (Meloidogyne incognita) Infection on Two African Indigenous Vegetables" Trends in Agricultural Sciences 3, no. 2: 100-106. https://doi.org/10.17311/tas.2024.100.106

This work is licensed under a Creative Commons Attribution 4.0 International License.